{"title":"找到一个良好的平衡:两个不同的染色质因子微调拟南芥幼苗的应激反应。","authors":"Rafal Archacki","doi":"10.1111/nph.19357","DOIUrl":null,"url":null,"abstract":"<p>Organisms maintain their integrity and achieve their goals of survival and reproduction by autonomously processing diverse inputs, such as environmental and intrinsic signals, into integrated responses; examples of this include initiating morphogenesis or changing the rate of growth. Plants, as sessile organisms, must continuously adapt to changing environmental conditions. Notable examples are water scarcity or salinity-induced stress faced by seeds or young seedlings, where the plants respond by delaying germination and by arresting postgermination development. These responses can be reversed upon the return of favorable conditions (Finch-Savage & Bassel, <span>2016</span>; Brandizzi, <span>2020</span>). While such plasticity offers advantages in nature, it is often undesirable in agricultural settings, as plants may also restrict their growth in response to moderate or temporary stress signals, which can lead to reductions in yield and harvest delays (Gupta <i>et al</i>., <span>2020</span>; Zhang <i>et al</i>., <span>2020</span>). Hence, unraveling the physiological and molecular basis of reaction to stress in <i>Arabidopsis thaliana</i> (Arabidopsis) and other plant species has been a major task in plant biology. It is now well established that stress responses are accompanied by changes in the activity of hundreds, or even thousands of genes, highlighting the remarkable ability of plants to adapt to conditions prevailing at a particular moment. These broad transcriptional responses are triggered and orchestrated by multiple signaling pathways comprising the action of hormones, transcription factors (TFs), and chromatin modifications (Takahashi <i>et al</i>., <span>2018</span>; Waadt <i>et al</i>., <span>2022</span>; Zhang <i>et al</i>., <span>2022</span>). Yet, despite significant progress, the emerging picture is still far from complete. In an article published in this issue of <i>New Phytologist</i>, Perrella <i>et al</i>. (<span>2023</span>; 166–179) report that two distinct chromatin factors, Histone Deacetylase Complex 1 (HDC1) and linker (H1) histones, are involved in shaping the proper transcriptional response to salt stress in Arabidopsis seedlings. They propose a ‘dual brake’ mechanism where HDC1 and H1 impact two different epigenetic marks to suppress some key stress-induced genes, thereby preventing hypersensitive responses.</p><p>In addition to its purely structural functions, chromatin is considered to be a major regulatory system that coordinates various genetic networks, including those involved in stress perception and response (Badeaux & Shi, <span>2013</span>). External and developmental cues can be transmitted through signaling cascades to chromatin-modifying enzymes, including histone acetyltransferases, methyltransferases, and many others (Bannister & Kouzarides, <span>2011</span>). The introduced modifications can directly or indirectly modulate chromatin structure, affecting its accessibility to different factors (e.g. TFs and polymerases) and, ultimately, gene activity. Transcriptionally active chromatin regions are typically marked by histone acetylation and trimethylation of histone H3 on lysine 4 (H3K4me3), while transcriptionally repressed regions are deacetylated and distinguished by trimethylation of histone H3 on lysine 9 (H3K9me3) or 27 (H3K27me3). Notably, these chromatin modifications can be removed by other enzymes, such as histone deacetylases (HDACs; Millán-Zambrano <i>et al</i>., <span>2022</span>). The main object of the study by Perrella <i>et al</i>. (<span>2023</span>), the HDC1 protein, has been shown to be a component of HDAC complexes (Perrella <i>et al</i>., <span>2013</span>; Mehdi <i>et al</i>., <span>2016</span>), and accordingly, the <i>hdc1</i> knockout mutation was reported to cause global hyperacetylation of H3 lysines 9 and 14 (H3K9K14; Perrella <i>et al</i>., <span>2013</span>).</p><p>Having previously established that <i>hdc1</i> mutant seedlings show a hypersensitive response to salt (Perrella <i>et al</i>., <span>2013</span>), the authors focused on the identification of gene targets regulated by HDC1 under these conditions. Surprisingly, despite the association of histone deacetylation with gene repression and increased global histone acetylation in the mutant, most of the differentially expressed genes (DEGs) found in salt-treated <i>hdc1-1</i> were downregulated. The authors hypothesized that a small number of direct HDC1 targets tune the sensitivity of stress perception in the seedlings, while most of the observed transcriptional differences are a consequence of enhanced stress sensitivity in the mutant. Therefore, they selected several candidate genes exhibiting H3K9K14 hyperacetylation and upregulation of the transcript in salt-treated <i>hdc1-1</i> as compared to control. Importantly, they provided genetic evidence in support of the key role of two of these genes by showing that knockout mutants of <i>LATE EMBRYOGENESIS-ABUNDANT</i> (<i>LEA</i>) or <i>MADS AFFECTING FLOWERING 5</i> (<i>MAF5</i>) both displayed lower sensitivity to salt treatment than wild-type plants and suppressed the salt-hypersensitive phenotype of <i>hdc1-1</i>. Thus, <i>LEA</i> and <i>MAF5</i> appear to mediate the stress-induced arrest of seedling development. It should be noted that while <i>LEA</i> encodes a member of a well-known protein family with roles in seed development (Candat <i>et al</i>., <span>2014</span>), until the study by Perrella <i>et al</i>. (<span>2023</span>) the known functions of MAF5 were limited to flowering control (Ratcliffe <i>et al</i>., <span>2003</span>). It has not been investigated whether two other genes, <i>ABSCISIC ACID INSENSITIVE 3</i> (ABI3) and <i>RAB GTPASE HOMOLOG B18</i> (RAB18), that show similar expression and H3K9/16Ac profiles to <i>LEA</i> and <i>MAF5</i> are also critical for the <i>hdc1</i> phenotype, and further studies await. Nonetheless, these findings highlight a recognized, but often neglected, possibility that the mutant phenotype can result from a small number of events (e.g. misregulation of few key genes), despite the wide changes in gene activity occurring in parallel.</p><p>Histone Deacetylase Complex 1 can interact with components of HDAC complexes as well as with several other proteins, including three variants of the linker histone H1 present in Arabidopsis (H1.1, H1.2, and H1.3; Rutowicz <i>et al</i>., <span>2015</span>; Perrella <i>et al</i>., <span>2016</span>). Perrella <i>et al</i>. (<span>2023</span>) revealed that the triple H1 mutation (<i>3h1</i>), similar to <i>hdc1</i>, confers sensitivity to salt. Furthermore, the transcript levels of the <i>ABI3</i>, <i>LEA</i>, <i>MAF5</i>, and <i>RAB18</i> genes showed a stronger induction upon salt treatment in <i>3h1</i> than in wild-type seedlings, although the levels of H3K9/16Ac were not altered. This suggests that HDC1-mediated deacetylation under salt stress is independent of H1 functions, but somehow H1 contributes to the establishment or maintenance of the proper expression levels of key genes. Therefore, the authors investigated other epigenetic marks that were previously reported to be affected in <i>h1</i> mutants (Zemach <i>et al</i>., <span>2013</span>; Rutowicz <i>et al</i>., <span>2019</span>). Remarkably, they found that in wild-type seedlings, salt treatment caused a strong increase in H3K27me3 levels in the studied genes, and both the <i>hdc1</i> and <i>h1</i> mutations prevented this effect. This indicates an additional role for HDC1 in facilitating stress-dependent deposition of H3K27me and suggests a functional link between HDC1 and H1. Linker histones have been shown to promote H3K27me3 enrichment at many genes in both animals and plants (Willcockson <i>et al</i>., <span>2021</span>; Teano <i>et al</i>., <span>2023</span>), and several other results support H1-HDC1 cooperation, including their direct interaction and nonadditive effects on transcript, as well as H3K9/16Ac and H3K27me3, levels under salt stress in the <i>h1.2 h1.3 hdc1</i> triple mutant. Unfortunately, the attempts of the authors to generate a full quadruple <i>3h1 hdc1</i> knockout were unsuccessful, likely because of its lethality, thereby implying that HDC1 and H1s have overlapping functions in development under normal conditions. Obviously, further studies will be needed to dissect the H1-HDC1 relationship at the molecular level and to resolve some outstanding questions. For example, based on their experiments using a truncated version of HDC1 which is capable of binding H1, the authors have suggested that H3 deacetylation is a prerequisite for H3K27me3 deposition upon salt stress. While this is likely in general, the mechanistic details remain unclear. Other questions that require clarification include: what serves as the primary signal for recruiting PRC2 complexes responsible for H3K27me3 deposition? Is it histone deacetylation itself, the presence of the HDC1-containing HDAC complex, H1, or a combination of these factors? Additionally, it will be important to determine whether H1 binding changes under stress conditions and whether it depends on H3K9/14 (de)acetylation and the presence of HDC1. Intriguingly, in both animals and plants, H1s have been functionally linked with a different histone acetylation mark, H3K56ac (Bernier <i>et al</i>., <span>2015</span>; Sheikh <i>et al</i>., <span>2023</span>). Therefore, it would be interesting to test whether HDC1 mediates deacetylation of this and/or other sites, or even H1, under salt stress. In the recent report by Sheikh <i>et al</i>. (<span>2023</span>), changes in H3K56ac levels were correlated with the role of Arabidopsis H1 in the regulation of the defense gene expression after pathogen challenge. However, H3K27me3 levels were not significantly altered in the <i>3h1</i> mutant under these conditions. Thus, the function of H1 in promoting deposition of H3K27me3 can be specific to gene and stress type. Finally, given that H1.3 represents a stress-inducible variant of H1 conserved in all angiosperms, the question of whether and how it contributes to the different H1-dependent stress responses remains to be resolved.</p><p>From a broader perspective, the findings reported by Perrella <i>et al</i>. (<span>2023</span>) extend our understanding of how plants moderate their developmental responses under adverse conditions. In the model proposed by the authors, HDC1 within the HDAC complex mediates the H3K9/14 deacetylation of the key stress-responsive genes <i>LEA</i> and <i>MAF5</i>, counteracting salt-induced acetylation, which is likely to prevent their excessive upregulation. Additionally, H3K27me3 levels in these genes increase with salt stress (Fig. 1). As the authors propose, the seemingly counterintuitive deposition of a repressive mark on stress-induced genes can act as a ‘dual brake’ to prevent overly sensitive responses. While other explanations are also possible, they would need experimental verification. For example, H3K27 methylation, known for its relative stability (Gallusci <i>et al</i>., <span>2023</span>), could potentially be deposited to restore efficient growth after the stress conditions disappear. It is also tempting to speculate that the increased H3K27me3 levels are important in limiting the activity of the selected genes following recurring stress. Otherwise, the genes could be primed for high expression, a process known to occur under different stress conditions (Bäurle, <span>2018</span>). However, this may be undesirable in some circumstances (Crisp <i>et al</i>., <span>2016</span>), as in the case of <i>LEA</i> and <i>MAF5</i> which were shown by Perrella <i>et al</i>. (<span>2023</span>) to promote growth arrest. Regardless of possible interpretations, the results of Perrella <i>et al</i>. (<span>2023</span>) may have important implications in the future, as reinforcing the dual brake mechanism could serve as an effective strategy to improve the performance of crop species experiencing moderate stresses in the field.</p>","PeriodicalId":214,"journal":{"name":"New Phytologist","volume":"241 1","pages":"7-9"},"PeriodicalIF":8.3000,"publicationDate":"2023-11-02","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://nph.onlinelibrary.wiley.com/doi/epdf/10.1111/nph.19357","citationCount":"0","resultStr":"{\"title\":\"Finding a good balance: two distinct chromatin factors fine-tune stress response in Arabidopsis seedlings\",\"authors\":\"Rafal Archacki\",\"doi\":\"10.1111/nph.19357\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p>Organisms maintain their integrity and achieve their goals of survival and reproduction by autonomously processing diverse inputs, such as environmental and intrinsic signals, into integrated responses; examples of this include initiating morphogenesis or changing the rate of growth. Plants, as sessile organisms, must continuously adapt to changing environmental conditions. Notable examples are water scarcity or salinity-induced stress faced by seeds or young seedlings, where the plants respond by delaying germination and by arresting postgermination development. These responses can be reversed upon the return of favorable conditions (Finch-Savage & Bassel, <span>2016</span>; Brandizzi, <span>2020</span>). While such plasticity offers advantages in nature, it is often undesirable in agricultural settings, as plants may also restrict their growth in response to moderate or temporary stress signals, which can lead to reductions in yield and harvest delays (Gupta <i>et al</i>., <span>2020</span>; Zhang <i>et al</i>., <span>2020</span>). Hence, unraveling the physiological and molecular basis of reaction to stress in <i>Arabidopsis thaliana</i> (Arabidopsis) and other plant species has been a major task in plant biology. It is now well established that stress responses are accompanied by changes in the activity of hundreds, or even thousands of genes, highlighting the remarkable ability of plants to adapt to conditions prevailing at a particular moment. These broad transcriptional responses are triggered and orchestrated by multiple signaling pathways comprising the action of hormones, transcription factors (TFs), and chromatin modifications (Takahashi <i>et al</i>., <span>2018</span>; Waadt <i>et al</i>., <span>2022</span>; Zhang <i>et al</i>., <span>2022</span>). Yet, despite significant progress, the emerging picture is still far from complete. In an article published in this issue of <i>New Phytologist</i>, Perrella <i>et al</i>. (<span>2023</span>; 166–179) report that two distinct chromatin factors, Histone Deacetylase Complex 1 (HDC1) and linker (H1) histones, are involved in shaping the proper transcriptional response to salt stress in Arabidopsis seedlings. They propose a ‘dual brake’ mechanism where HDC1 and H1 impact two different epigenetic marks to suppress some key stress-induced genes, thereby preventing hypersensitive responses.</p><p>In addition to its purely structural functions, chromatin is considered to be a major regulatory system that coordinates various genetic networks, including those involved in stress perception and response (Badeaux & Shi, <span>2013</span>). External and developmental cues can be transmitted through signaling cascades to chromatin-modifying enzymes, including histone acetyltransferases, methyltransferases, and many others (Bannister & Kouzarides, <span>2011</span>). The introduced modifications can directly or indirectly modulate chromatin structure, affecting its accessibility to different factors (e.g. TFs and polymerases) and, ultimately, gene activity. Transcriptionally active chromatin regions are typically marked by histone acetylation and trimethylation of histone H3 on lysine 4 (H3K4me3), while transcriptionally repressed regions are deacetylated and distinguished by trimethylation of histone H3 on lysine 9 (H3K9me3) or 27 (H3K27me3). Notably, these chromatin modifications can be removed by other enzymes, such as histone deacetylases (HDACs; Millán-Zambrano <i>et al</i>., <span>2022</span>). The main object of the study by Perrella <i>et al</i>. (<span>2023</span>), the HDC1 protein, has been shown to be a component of HDAC complexes (Perrella <i>et al</i>., <span>2013</span>; Mehdi <i>et al</i>., <span>2016</span>), and accordingly, the <i>hdc1</i> knockout mutation was reported to cause global hyperacetylation of H3 lysines 9 and 14 (H3K9K14; Perrella <i>et al</i>., <span>2013</span>).</p><p>Having previously established that <i>hdc1</i> mutant seedlings show a hypersensitive response to salt (Perrella <i>et al</i>., <span>2013</span>), the authors focused on the identification of gene targets regulated by HDC1 under these conditions. Surprisingly, despite the association of histone deacetylation with gene repression and increased global histone acetylation in the mutant, most of the differentially expressed genes (DEGs) found in salt-treated <i>hdc1-1</i> were downregulated. The authors hypothesized that a small number of direct HDC1 targets tune the sensitivity of stress perception in the seedlings, while most of the observed transcriptional differences are a consequence of enhanced stress sensitivity in the mutant. Therefore, they selected several candidate genes exhibiting H3K9K14 hyperacetylation and upregulation of the transcript in salt-treated <i>hdc1-1</i> as compared to control. Importantly, they provided genetic evidence in support of the key role of two of these genes by showing that knockout mutants of <i>LATE EMBRYOGENESIS-ABUNDANT</i> (<i>LEA</i>) or <i>MADS AFFECTING FLOWERING 5</i> (<i>MAF5</i>) both displayed lower sensitivity to salt treatment than wild-type plants and suppressed the salt-hypersensitive phenotype of <i>hdc1-1</i>. Thus, <i>LEA</i> and <i>MAF5</i> appear to mediate the stress-induced arrest of seedling development. It should be noted that while <i>LEA</i> encodes a member of a well-known protein family with roles in seed development (Candat <i>et al</i>., <span>2014</span>), until the study by Perrella <i>et al</i>. (<span>2023</span>) the known functions of MAF5 were limited to flowering control (Ratcliffe <i>et al</i>., <span>2003</span>). It has not been investigated whether two other genes, <i>ABSCISIC ACID INSENSITIVE 3</i> (ABI3) and <i>RAB GTPASE HOMOLOG B18</i> (RAB18), that show similar expression and H3K9/16Ac profiles to <i>LEA</i> and <i>MAF5</i> are also critical for the <i>hdc1</i> phenotype, and further studies await. Nonetheless, these findings highlight a recognized, but often neglected, possibility that the mutant phenotype can result from a small number of events (e.g. misregulation of few key genes), despite the wide changes in gene activity occurring in parallel.</p><p>Histone Deacetylase Complex 1 can interact with components of HDAC complexes as well as with several other proteins, including three variants of the linker histone H1 present in Arabidopsis (H1.1, H1.2, and H1.3; Rutowicz <i>et al</i>., <span>2015</span>; Perrella <i>et al</i>., <span>2016</span>). Perrella <i>et al</i>. (<span>2023</span>) revealed that the triple H1 mutation (<i>3h1</i>), similar to <i>hdc1</i>, confers sensitivity to salt. Furthermore, the transcript levels of the <i>ABI3</i>, <i>LEA</i>, <i>MAF5</i>, and <i>RAB18</i> genes showed a stronger induction upon salt treatment in <i>3h1</i> than in wild-type seedlings, although the levels of H3K9/16Ac were not altered. This suggests that HDC1-mediated deacetylation under salt stress is independent of H1 functions, but somehow H1 contributes to the establishment or maintenance of the proper expression levels of key genes. Therefore, the authors investigated other epigenetic marks that were previously reported to be affected in <i>h1</i> mutants (Zemach <i>et al</i>., <span>2013</span>; Rutowicz <i>et al</i>., <span>2019</span>). Remarkably, they found that in wild-type seedlings, salt treatment caused a strong increase in H3K27me3 levels in the studied genes, and both the <i>hdc1</i> and <i>h1</i> mutations prevented this effect. This indicates an additional role for HDC1 in facilitating stress-dependent deposition of H3K27me and suggests a functional link between HDC1 and H1. Linker histones have been shown to promote H3K27me3 enrichment at many genes in both animals and plants (Willcockson <i>et al</i>., <span>2021</span>; Teano <i>et al</i>., <span>2023</span>), and several other results support H1-HDC1 cooperation, including their direct interaction and nonadditive effects on transcript, as well as H3K9/16Ac and H3K27me3, levels under salt stress in the <i>h1.2 h1.3 hdc1</i> triple mutant. Unfortunately, the attempts of the authors to generate a full quadruple <i>3h1 hdc1</i> knockout were unsuccessful, likely because of its lethality, thereby implying that HDC1 and H1s have overlapping functions in development under normal conditions. Obviously, further studies will be needed to dissect the H1-HDC1 relationship at the molecular level and to resolve some outstanding questions. For example, based on their experiments using a truncated version of HDC1 which is capable of binding H1, the authors have suggested that H3 deacetylation is a prerequisite for H3K27me3 deposition upon salt stress. While this is likely in general, the mechanistic details remain unclear. Other questions that require clarification include: what serves as the primary signal for recruiting PRC2 complexes responsible for H3K27me3 deposition? Is it histone deacetylation itself, the presence of the HDC1-containing HDAC complex, H1, or a combination of these factors? Additionally, it will be important to determine whether H1 binding changes under stress conditions and whether it depends on H3K9/14 (de)acetylation and the presence of HDC1. Intriguingly, in both animals and plants, H1s have been functionally linked with a different histone acetylation mark, H3K56ac (Bernier <i>et al</i>., <span>2015</span>; Sheikh <i>et al</i>., <span>2023</span>). Therefore, it would be interesting to test whether HDC1 mediates deacetylation of this and/or other sites, or even H1, under salt stress. In the recent report by Sheikh <i>et al</i>. (<span>2023</span>), changes in H3K56ac levels were correlated with the role of Arabidopsis H1 in the regulation of the defense gene expression after pathogen challenge. However, H3K27me3 levels were not significantly altered in the <i>3h1</i> mutant under these conditions. Thus, the function of H1 in promoting deposition of H3K27me3 can be specific to gene and stress type. Finally, given that H1.3 represents a stress-inducible variant of H1 conserved in all angiosperms, the question of whether and how it contributes to the different H1-dependent stress responses remains to be resolved.</p><p>From a broader perspective, the findings reported by Perrella <i>et al</i>. (<span>2023</span>) extend our understanding of how plants moderate their developmental responses under adverse conditions. In the model proposed by the authors, HDC1 within the HDAC complex mediates the H3K9/14 deacetylation of the key stress-responsive genes <i>LEA</i> and <i>MAF5</i>, counteracting salt-induced acetylation, which is likely to prevent their excessive upregulation. Additionally, H3K27me3 levels in these genes increase with salt stress (Fig. 1). As the authors propose, the seemingly counterintuitive deposition of a repressive mark on stress-induced genes can act as a ‘dual brake’ to prevent overly sensitive responses. While other explanations are also possible, they would need experimental verification. For example, H3K27 methylation, known for its relative stability (Gallusci <i>et al</i>., <span>2023</span>), could potentially be deposited to restore efficient growth after the stress conditions disappear. It is also tempting to speculate that the increased H3K27me3 levels are important in limiting the activity of the selected genes following recurring stress. Otherwise, the genes could be primed for high expression, a process known to occur under different stress conditions (Bäurle, <span>2018</span>). However, this may be undesirable in some circumstances (Crisp <i>et al</i>., <span>2016</span>), as in the case of <i>LEA</i> and <i>MAF5</i> which were shown by Perrella <i>et al</i>. (<span>2023</span>) to promote growth arrest. Regardless of possible interpretations, the results of Perrella <i>et al</i>. (<span>2023</span>) may have important implications in the future, as reinforcing the dual brake mechanism could serve as an effective strategy to improve the performance of crop species experiencing moderate stresses in the field.</p>\",\"PeriodicalId\":214,\"journal\":{\"name\":\"New Phytologist\",\"volume\":\"241 1\",\"pages\":\"7-9\"},\"PeriodicalIF\":8.3000,\"publicationDate\":\"2023-11-02\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://nph.onlinelibrary.wiley.com/doi/epdf/10.1111/nph.19357\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"New Phytologist\",\"FirstCategoryId\":\"99\",\"ListUrlMain\":\"https://onlinelibrary.wiley.com/doi/10.1111/nph.19357\",\"RegionNum\":1,\"RegionCategory\":\"生物学\",\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"Q1\",\"JCRName\":\"PLANT SCIENCES\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"New Phytologist","FirstCategoryId":"99","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1111/nph.19357","RegionNum":1,"RegionCategory":"生物学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"PLANT SCIENCES","Score":null,"Total":0}
引用次数: 0
摘要
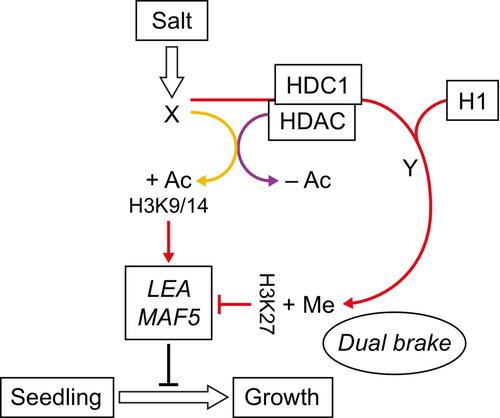
生物体通过自主地将各种输入(如环境和内在信号)加工成综合反应来维持其完整性并实现其生存和繁殖目标;这方面的例子包括启动形态发生或改变生长速度。植物作为无根生物,必须不断适应不断变化的环境条件。值得注意的例子是种子或幼苗面临的缺水或盐碱胁迫,植物的反应是延迟发芽和阻止发芽后的发育。在有利条件回归时,这些反应可能会被逆转(芬奇-萨维奇&;巴塞尔因车祸,2016;Brandizzi, 2020)。虽然这种可塑性在自然界中具有优势,但在农业环境中往往是不可取的,因为植物也可能对中等或暂时的应激信号作出反应,限制其生长,这可能导致产量减少和收获延迟(Gupta等人,2020;Zhang等人,2020)。因此,揭示拟南芥(Arabidopsis thaliana,简称拟南芥)等植物对胁迫反应的生理和分子基础一直是植物生物学研究的重要课题。现在已经确定的是,应激反应伴随着数百甚至数千个基因活动的变化,这突出了植物适应特定时刻普遍存在的条件的非凡能力。这些广泛的转录反应由多种信号通路触发和协调,包括激素、转录因子(tf)和染色质修饰的作用(Takahashi等人,2018;Waadt et al., 2022;Zhang等人,2022)。然而,尽管取得了重大进展,但新出现的情况仍远未完成。在新植物学家发表的一篇文章中,Perrella et al. (2023;166-179)报道了两种不同的染色质因子,组蛋白去乙酰化酶复合体1 (HDC1)和连接体(H1)组蛋白,参与了拟南芥幼苗对盐胁迫的转录反应。他们提出了一种“双刹车”机制,HDC1和H1影响两种不同的表观遗传标记,抑制一些关键的应激诱导基因,从而防止过敏反应。除了其纯粹的结构功能,染色质被认为是一个主要的调节系统,协调各种遗传网络,包括那些参与应激感知和反应(Badeaux &史,2013)。外部和发育线索可以通过信号级联传递到染色质修饰酶,包括组蛋白乙酰转移酶、甲基转移酶和许多其他酶(Bannister &Kouzarides, 2011)。引入的修饰可以直接或间接地调节染色质结构,影响其对不同因子(如tf和聚合酶)的可及性,并最终影响基因活性。转录活性染色质区域通常以赖氨酸4上的组蛋白H3乙酰化和三甲基化(H3K4me3)为标志,而转录抑制区域则以赖氨酸9 (H3K9me3)或27 (H3K27me3)上的组蛋白H3去乙酰化和三甲基化来区分。值得注意的是,这些染色质修饰可以被其他酶去除,如组蛋白去乙酰化酶(hdac;Millán-Zambrano et al., 2022)。Perrella et al.(2023)研究的主要对象HDC1蛋白已被证明是HDAC复合物的一个组成部分(Perrella et al., 2013;Mehdi等人,2016),因此,据报道hdc1基因敲除突变导致H3赖氨酸9和14的整体超乙酰化(H3K9K14;Perrella et al., 2013)。先前已经确定hdc1突变苗对盐表现出过敏反应(Perrella et al., 2013),作者专注于在这些条件下鉴定由hdc1调控的基因靶点。令人惊讶的是,尽管在突变体中组蛋白去乙酰化与基因抑制和全局组蛋白乙酰化增加有关,但在盐处理的hdc1-1中发现的大多数差异表达基因(DEGs)被下调。作者假设,少量直接的HDC1靶点调节了幼苗对压力感知的敏感性,而大多数观察到的转录差异是突变体对压力敏感性增强的结果。因此,他们选择了几个候选基因,与对照相比,在盐处理的hdc1-1中表现出H3K9K14超乙酰化和转录上调。重要的是,他们提供了支持其中两个基因的关键作用的遗传证据,表明LATE embryogenetic - abundant (LEA)或MADS影响开花5 (MAF5)的敲除突变体对盐处理的敏感性都低于野生型植物,并抑制hdc1-1的盐敏感表型。因此,LEA和MAF5似乎介导了胁迫诱导的幼苗发育停滞。 值得注意的是,LEA编码的是一个众所周知的在种子发育中起作用的蛋白家族成员(Candat et al., 2014),在Perrella et al.(2023)的研究之前,已知的MAF5功能仅限于控制开花(Ratcliffe et al., 2003)。目前尚未研究另外两个与LEA和MAF5表达和H3K9/16Ac谱相似的基因ABSCISIC ACID INSENSITIVE 3 (ABI3)和RAB GTPASE HOMOLOG B18 (RAB18)是否也是hdc1表型的关键基因,有待进一步的研究。尽管如此,这些发现强调了一个公认但经常被忽视的可能性,即突变表型可能是由少数事件(例如,少数关键基因的错误调节)引起的,尽管基因活性的广泛变化同时发生。组蛋白去乙酰化酶复合体1可以与HDAC复合体的组分以及其他几种蛋白质相互作用,包括拟南芥中存在的连接组蛋白H1的三种变体(H1.1, H1.2和H1.3;Rutowicz et al., 2015;Perrella et al., 2016)。Perrella等人(2023)发现,三H1突变(3h1)与hdc1相似,赋予盐敏感性。此外,尽管H3K9/16Ac的水平没有改变,但3h1中ABI3、LEA、MAF5和RAB18基因的转录水平在盐处理后的诱导作用强于野生型幼苗。这表明盐胁迫下hdc1介导的去乙酰化与H1的功能无关,但H1在某种程度上有助于关键基因适当表达水平的建立或维持。因此,作者研究了先前报道的在h1突变体中受影响的其他表观遗传标记(Zemach et al., 2013;Rutowicz et al., 2019)。值得注意的是,他们发现,在野生型幼苗中,盐处理导致研究基因中H3K27me3水平的强烈增加,而hdc1和h1突变都阻止了这种影响。这表明HDC1在促进H3K27me的应力依赖性沉积中起着额外的作用,并表明HDC1和H1之间存在功能联系。在动物和植物中,连接子组蛋白已被证明可促进H3K27me3在许多基因上的富集(Willcockson等人,2021;Teano et al., 2023)和其他几个结果支持H1-HDC1合作,包括它们在盐胁迫下对转录本以及H3K9/16Ac和H3K27me3水平的直接相互作用和非加性作用。不幸的是,作者试图产生一个完整的四重3h1 hdc1敲除没有成功,可能是因为它的致命性,从而暗示hdc1和H1s在正常条件下的发育中具有重叠的功能。显然,需要进一步的研究在分子水平上剖析H1-HDC1的关系,并解决一些悬而未决的问题。例如,基于他们使用能够结合H1的HDC1的截断版本的实验,作者提出H3去乙酰化是H3K27me3在盐胁迫下沉积的先决条件。虽然这在总体上是可能的,但机制细节仍不清楚。其他需要澄清的问题包括:募集负责H3K27me3沉积的PRC2复合物的主要信号是什么?是组蛋白去乙酰化本身,含有hdc1的HDAC复合体H1的存在,还是这些因素的组合?此外,确定H1结合是否在应激条件下发生变化以及它是否依赖于H3K9/14(去)乙酰化和HDC1的存在也很重要。有趣的是,在动物和植物中,H1s都与不同的组蛋白乙酰化标记H3K56ac有功能联系(Bernier et al., 2015;Sheikh et al., 2023)。因此,测试HDC1是否在盐胁迫下介导该位点和/或其他位点,甚至H1的去乙酰化将是有趣的。在Sheikh等人(2023)最近的报道中,H3K56ac水平的变化与拟南芥H1在病原体攻击后调控防御基因表达的作用相关。然而,在这些条件下,H3K27me3水平在3h1突变体中没有显著改变。因此,H1促进H3K27me3沉积的功能可能与基因和应激类型有关。最后,考虑到H1.3代表了在所有被子植物中保守的应激诱导的H1变异,它是否以及如何促进不同的H1依赖的应激反应的问题仍有待解决。从更广泛的角度来看,Perrella等人(2023)报告的研究结果扩展了我们对植物如何在不利条件下调节其发育反应的理解。在作者提出的模型中,HDAC复合体中的HDC1介导关键应激反应基因LEA和MAF5的H3K9/14去乙酰化,抵消盐诱导的乙酰化,这可能会阻止它们的过度上调。 此外,这些基因中的H3K27me3水平随着盐胁迫而增加(图1)。正如作者所提出的,应激诱导基因上抑制标记的沉积看似违反直觉,可以起到“双重刹车”的作用,防止过度敏感的反应。虽然其他解释也有可能,但它们需要实验验证。例如,以其相对稳定性而闻名的H3K27甲基化(Gallusci et al., 2023)可能在应激条件消失后沉积以恢复有效生长。这也很容易让人推测,H3K27me3水平的增加在限制反复出现的压力后选定基因的活性方面是重要的。否则,这些基因可能会引发高表达,这一过程已知会在不同的应激条件下发生(Bäurle, 2018)。然而,在某些情况下,这可能是不可取的(Crisp et al., 2016),例如Perrella et al.(2023)表明LEA和MAF5会促进生长停滞。不管可能的解释是什么,Perrella等人(2023)的结果可能对未来具有重要意义,因为加强双制动机制可以作为一种有效的策略来改善田间遭受中度胁迫的作物品种的生产性能。
Finding a good balance: two distinct chromatin factors fine-tune stress response in Arabidopsis seedlings
Organisms maintain their integrity and achieve their goals of survival and reproduction by autonomously processing diverse inputs, such as environmental and intrinsic signals, into integrated responses; examples of this include initiating morphogenesis or changing the rate of growth. Plants, as sessile organisms, must continuously adapt to changing environmental conditions. Notable examples are water scarcity or salinity-induced stress faced by seeds or young seedlings, where the plants respond by delaying germination and by arresting postgermination development. These responses can be reversed upon the return of favorable conditions (Finch-Savage & Bassel, 2016; Brandizzi, 2020). While such plasticity offers advantages in nature, it is often undesirable in agricultural settings, as plants may also restrict their growth in response to moderate or temporary stress signals, which can lead to reductions in yield and harvest delays (Gupta et al., 2020; Zhang et al., 2020). Hence, unraveling the physiological and molecular basis of reaction to stress in Arabidopsis thaliana (Arabidopsis) and other plant species has been a major task in plant biology. It is now well established that stress responses are accompanied by changes in the activity of hundreds, or even thousands of genes, highlighting the remarkable ability of plants to adapt to conditions prevailing at a particular moment. These broad transcriptional responses are triggered and orchestrated by multiple signaling pathways comprising the action of hormones, transcription factors (TFs), and chromatin modifications (Takahashi et al., 2018; Waadt et al., 2022; Zhang et al., 2022). Yet, despite significant progress, the emerging picture is still far from complete. In an article published in this issue of New Phytologist, Perrella et al. (2023; 166–179) report that two distinct chromatin factors, Histone Deacetylase Complex 1 (HDC1) and linker (H1) histones, are involved in shaping the proper transcriptional response to salt stress in Arabidopsis seedlings. They propose a ‘dual brake’ mechanism where HDC1 and H1 impact two different epigenetic marks to suppress some key stress-induced genes, thereby preventing hypersensitive responses.
In addition to its purely structural functions, chromatin is considered to be a major regulatory system that coordinates various genetic networks, including those involved in stress perception and response (Badeaux & Shi, 2013). External and developmental cues can be transmitted through signaling cascades to chromatin-modifying enzymes, including histone acetyltransferases, methyltransferases, and many others (Bannister & Kouzarides, 2011). The introduced modifications can directly or indirectly modulate chromatin structure, affecting its accessibility to different factors (e.g. TFs and polymerases) and, ultimately, gene activity. Transcriptionally active chromatin regions are typically marked by histone acetylation and trimethylation of histone H3 on lysine 4 (H3K4me3), while transcriptionally repressed regions are deacetylated and distinguished by trimethylation of histone H3 on lysine 9 (H3K9me3) or 27 (H3K27me3). Notably, these chromatin modifications can be removed by other enzymes, such as histone deacetylases (HDACs; Millán-Zambrano et al., 2022). The main object of the study by Perrella et al. (2023), the HDC1 protein, has been shown to be a component of HDAC complexes (Perrella et al., 2013; Mehdi et al., 2016), and accordingly, the hdc1 knockout mutation was reported to cause global hyperacetylation of H3 lysines 9 and 14 (H3K9K14; Perrella et al., 2013).
Having previously established that hdc1 mutant seedlings show a hypersensitive response to salt (Perrella et al., 2013), the authors focused on the identification of gene targets regulated by HDC1 under these conditions. Surprisingly, despite the association of histone deacetylation with gene repression and increased global histone acetylation in the mutant, most of the differentially expressed genes (DEGs) found in salt-treated hdc1-1 were downregulated. The authors hypothesized that a small number of direct HDC1 targets tune the sensitivity of stress perception in the seedlings, while most of the observed transcriptional differences are a consequence of enhanced stress sensitivity in the mutant. Therefore, they selected several candidate genes exhibiting H3K9K14 hyperacetylation and upregulation of the transcript in salt-treated hdc1-1 as compared to control. Importantly, they provided genetic evidence in support of the key role of two of these genes by showing that knockout mutants of LATE EMBRYOGENESIS-ABUNDANT (LEA) or MADS AFFECTING FLOWERING 5 (MAF5) both displayed lower sensitivity to salt treatment than wild-type plants and suppressed the salt-hypersensitive phenotype of hdc1-1. Thus, LEA and MAF5 appear to mediate the stress-induced arrest of seedling development. It should be noted that while LEA encodes a member of a well-known protein family with roles in seed development (Candat et al., 2014), until the study by Perrella et al. (2023) the known functions of MAF5 were limited to flowering control (Ratcliffe et al., 2003). It has not been investigated whether two other genes, ABSCISIC ACID INSENSITIVE 3 (ABI3) and RAB GTPASE HOMOLOG B18 (RAB18), that show similar expression and H3K9/16Ac profiles to LEA and MAF5 are also critical for the hdc1 phenotype, and further studies await. Nonetheless, these findings highlight a recognized, but often neglected, possibility that the mutant phenotype can result from a small number of events (e.g. misregulation of few key genes), despite the wide changes in gene activity occurring in parallel.
Histone Deacetylase Complex 1 can interact with components of HDAC complexes as well as with several other proteins, including three variants of the linker histone H1 present in Arabidopsis (H1.1, H1.2, and H1.3; Rutowicz et al., 2015; Perrella et al., 2016). Perrella et al. (2023) revealed that the triple H1 mutation (3h1), similar to hdc1, confers sensitivity to salt. Furthermore, the transcript levels of the ABI3, LEA, MAF5, and RAB18 genes showed a stronger induction upon salt treatment in 3h1 than in wild-type seedlings, although the levels of H3K9/16Ac were not altered. This suggests that HDC1-mediated deacetylation under salt stress is independent of H1 functions, but somehow H1 contributes to the establishment or maintenance of the proper expression levels of key genes. Therefore, the authors investigated other epigenetic marks that were previously reported to be affected in h1 mutants (Zemach et al., 2013; Rutowicz et al., 2019). Remarkably, they found that in wild-type seedlings, salt treatment caused a strong increase in H3K27me3 levels in the studied genes, and both the hdc1 and h1 mutations prevented this effect. This indicates an additional role for HDC1 in facilitating stress-dependent deposition of H3K27me and suggests a functional link between HDC1 and H1. Linker histones have been shown to promote H3K27me3 enrichment at many genes in both animals and plants (Willcockson et al., 2021; Teano et al., 2023), and several other results support H1-HDC1 cooperation, including their direct interaction and nonadditive effects on transcript, as well as H3K9/16Ac and H3K27me3, levels under salt stress in the h1.2 h1.3 hdc1 triple mutant. Unfortunately, the attempts of the authors to generate a full quadruple 3h1 hdc1 knockout were unsuccessful, likely because of its lethality, thereby implying that HDC1 and H1s have overlapping functions in development under normal conditions. Obviously, further studies will be needed to dissect the H1-HDC1 relationship at the molecular level and to resolve some outstanding questions. For example, based on their experiments using a truncated version of HDC1 which is capable of binding H1, the authors have suggested that H3 deacetylation is a prerequisite for H3K27me3 deposition upon salt stress. While this is likely in general, the mechanistic details remain unclear. Other questions that require clarification include: what serves as the primary signal for recruiting PRC2 complexes responsible for H3K27me3 deposition? Is it histone deacetylation itself, the presence of the HDC1-containing HDAC complex, H1, or a combination of these factors? Additionally, it will be important to determine whether H1 binding changes under stress conditions and whether it depends on H3K9/14 (de)acetylation and the presence of HDC1. Intriguingly, in both animals and plants, H1s have been functionally linked with a different histone acetylation mark, H3K56ac (Bernier et al., 2015; Sheikh et al., 2023). Therefore, it would be interesting to test whether HDC1 mediates deacetylation of this and/or other sites, or even H1, under salt stress. In the recent report by Sheikh et al. (2023), changes in H3K56ac levels were correlated with the role of Arabidopsis H1 in the regulation of the defense gene expression after pathogen challenge. However, H3K27me3 levels were not significantly altered in the 3h1 mutant under these conditions. Thus, the function of H1 in promoting deposition of H3K27me3 can be specific to gene and stress type. Finally, given that H1.3 represents a stress-inducible variant of H1 conserved in all angiosperms, the question of whether and how it contributes to the different H1-dependent stress responses remains to be resolved.
From a broader perspective, the findings reported by Perrella et al. (2023) extend our understanding of how plants moderate their developmental responses under adverse conditions. In the model proposed by the authors, HDC1 within the HDAC complex mediates the H3K9/14 deacetylation of the key stress-responsive genes LEA and MAF5, counteracting salt-induced acetylation, which is likely to prevent their excessive upregulation. Additionally, H3K27me3 levels in these genes increase with salt stress (Fig. 1). As the authors propose, the seemingly counterintuitive deposition of a repressive mark on stress-induced genes can act as a ‘dual brake’ to prevent overly sensitive responses. While other explanations are also possible, they would need experimental verification. For example, H3K27 methylation, known for its relative stability (Gallusci et al., 2023), could potentially be deposited to restore efficient growth after the stress conditions disappear. It is also tempting to speculate that the increased H3K27me3 levels are important in limiting the activity of the selected genes following recurring stress. Otherwise, the genes could be primed for high expression, a process known to occur under different stress conditions (Bäurle, 2018). However, this may be undesirable in some circumstances (Crisp et al., 2016), as in the case of LEA and MAF5 which were shown by Perrella et al. (2023) to promote growth arrest. Regardless of possible interpretations, the results of Perrella et al. (2023) may have important implications in the future, as reinforcing the dual brake mechanism could serve as an effective strategy to improve the performance of crop species experiencing moderate stresses in the field.
期刊介绍:
New Phytologist is an international electronic journal published 24 times a year. It is owned by the New Phytologist Foundation, a non-profit-making charitable organization dedicated to promoting plant science. The journal publishes excellent, novel, rigorous, and timely research and scholarship in plant science and its applications. The articles cover topics in five sections: Physiology & Development, Environment, Interaction, Evolution, and Transformative Plant Biotechnology. These sections encompass intracellular processes, global environmental change, and encourage cross-disciplinary approaches. The journal recognizes the use of techniques from molecular and cell biology, functional genomics, modeling, and system-based approaches in plant science. Abstracting and Indexing Information for New Phytologist includes Academic Search, AgBiotech News & Information, Agroforestry Abstracts, Biochemistry & Biophysics Citation Index, Botanical Pesticides, CAB Abstracts®, Environment Index, Global Health, and Plant Breeding Abstracts, and others.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


