Stephanie E. Hampton, Jill S. Baron, Robert Ladwig, Ryan P. McClure, Michael F. Meyer, Isabella A. Oleksy, Anna Shampain
{"title":"气候变暖引起的底栖氧化还原变化是高海拔湖泊底栖藻类水华加剧的潜在驱动因素","authors":"Stephanie E. Hampton, Jill S. Baron, Robert Ladwig, Ryan P. McClure, Michael F. Meyer, Isabella A. Oleksy, Anna Shampain","doi":"10.1002/lol2.10357","DOIUrl":null,"url":null,"abstract":"<p>Blooms of algae attached to the bottom of nearshore environments have increasingly been observed in clear, nutrient-poor lakes worldwide (Vadeboncoeur et al. <span>2021</span>). The phenomenon is puzzling for at least two reasons. First, such nearshore benthic blooms appear to be common across heterogeneous contexts—from the world's largest lake in Siberia to small mountain lakes in the Rocky Mountains of North America. Second, the blooms are enigmatic because they have been observed in the near absence of direct human disturbances, such as those in well-protected, remote mountain lakes. A recent review (Vadeboncoeur et al. <span>2021</span>) highlights the diversity of mechanisms through which these nearshore benthic blooms may be triggered—such as nutrient loading through atmospheric deposition or human activity within the watershed, changes in hydrodynamics associated with climate change, and food web alterations that reduce grazing pressure. Yet none of these potential drivers are common across the lakes where nearshore benthic algal blooms have been reported. Here, we suggest that a previously unappreciated driver is common across all lakes that experience warming associated with climate change—elevated nearshore temperatures that reduce oxygen and promote nutrient release from sediments under the biofilm. In principle, this phenomenon should be particularly notable in high-elevation lakes because water at higher elevation holds less oxygen at saturation, high-elevation lakes are warming more quickly than their lowland counterparts, and they can receive additional warming from relatively high solar radiation (Fig. 1). We briefly review the evidence in support of this concept, recognizing that the data necessary to rigorously test it do not yet exist; however, such data collection is feasible.</p><p>Algal taxa in oligotrophic systems are well adapted to naturally high-light and nutrient-poor conditions (Cantonati and Lowe <span>2014</span>). Because nutrients at the sediment–water interface can be rapidly cycled, nutrient concentrations in the water column may not be reliable indicators of nutrient availability for benthic primary producers (Vadeboncoeur and Power <span>2017</span>). Hot spots or hot moments of elevated nutrient concentrations in littoral regions favor green algae, leading to thick filamentous mats (also called filamentous algal blooms; Oleksy et al. <span>2021</span>; Vadeboncoeur et al. <span>2021</span>). These periphyton blooms appear to be an emergent response to changing environmental conditions in oligotrophic systems, yet the ultimate drivers behind their increased occurrence are debated and often obfuscated by confounding processes. For example, in Lake Baikal, increasing evidence has pointed to highly localized wastewater inputs leading to increased <i>Ulothrix</i> spp. and <i>Spirogyra</i> spp. abundance (Timoshkin et al. <span>2018</span>; Meyer et al. <span>2022</span>). Similarly, Lake Tahoe has experienced increased periphyton biomass likely due to a mix of wastewater-associated eutrophication, groundwater leachate, and complex physical and hydrological processes (Naranjo et al. <span>2019</span>; Atkins et al. <span>2022</span>). Although observations of increased periphyton blooms in lakes with adjacent human population can draw links between human activity and in-lake periphyton responses (e.g., Rosenberger et al. <span>2008</span>), these blooms also appear to be increasing in remote, high-elevation systems that lack permanent human settlements (Oleksy et al. <span>2020</span>).</p><p>High-elevation lakes located in mountainous regions are frequently characterized by low-nutrient concentrations, long periods of ice cover, high solar and UV radiation, as well as heterogeneous landscapes associated with complex topography (Moser et al. <span>2019</span>). Despite their remote locations, historically oligotrophic high-elevation lakes have experienced primary productivity increases that sometimes have been attributed to global change, including atmospheric deposition of reactive nitrogen and phosphorus, species introductions, and climate change (Ballantyne et al. <span>2011</span>; Saros et al. <span>2011</span>; Catalan et al. <span>2013</span>; Brahney et al. <span>2014</span>; Sadro et al. <span>2018</span>; Vadeboncoeur et al. <span>2021</span>). Most of this literature describes changes to water column productivity. However, numerous studies have shown that the frequently neglected benthic habitats of oligotrophic lakes, including those at high elevations, dominate ecosystem processes of primary production and respiration (reviewed by Vadeboncoeur and Power <span>2017</span>). Changes in benthic algal assemblages previously may have been overlooked. For instance, within lakes in the southern Rocky Mountains of North America, sediment records indicate that nearshore algal assemblages have changed appreciably in recent decades, with 200–300% increases in periphytic green algal species (Oleksy et al. <span>2020</span>).</p><p>Although sources and magnitude of nutrient inputs vary greatly across lakes worldwide, a common factor is warming associated with climate change (O'Reilly et al. <span>2015</span>), which can promote oxygen depletion (Jane et al. <span>2021</span>). Mountainous regions are warming faster than adjacent low-lying areas (Pepin et al. <span>2022</span>), and warm water holds less oxygen than cold water. In a high-elevation lake, relative to its lower elevation counterparts, the water column holds lower oxygen at saturation due to the inverse relationship between elevation (atmospheric pressure) and the oxygen capacity of water (Fig. 1). Hutchinson (<span>1957</span>) briefly alluded to the potential for this elevation–oxygen relationship to alter benthic ecology. The importance of not only oxygen content but also partial pressure is increasingly recognized as affecting biodiversity patterns among aquatic invertebrates (e.g., Verberk et al. <span>2011</span>). Our mass balance calculation using a vertical 1D reaction–diffusion model suggests that elevation effects on oxygen in a warming mountain lake could account for sediment anoxia occurring days to weeks earlier than would occur in a warming lake at sea level (Supporting Information). High solar radiation in mountain lakes can contribute to additional warming, and nearshore temperatures can increase markedly relative to the offshore. In some cases, as nearshore temperatures warm, a thermal bar may develop, reducing mixing and allowing littoral water temperatures to become even warmer (Vadeboncoeur et al. <span>2021</span>). Warming sediments can quickly stimulate heterotrophic microbial processes that consume oxygen (Bergström et al. <span>2010</span>). Convective cooling in the littoral zones can further limit horizontal exchange of oxygen and nutrients with offshore waters (Doda et al. <span>2023</span>). These combined factors could explain why mountain lake nearshore blooms have been so noticeable in the absence of other clear drivers.</p><p>Warming littoral zone temperatures can decrease oxygen available for aerobic metabolism, creating an environment more favorable to anaerobic metabolism just a few millimeters into the sediments (Fig. 1, Schlesinger and Bernhardt <span>2013</span>), particularly at night when photosynthesis does not produce oxygen. Microbial consumption of oxygen in sediments frequently exceeds the oxygen supply through diffusion (Fenchel and Finlay <span>2008</span>). Without oxygen, microbes cannot use oxidative phosphorylation to convert organic matter into CO<sub>2</sub>, and instead must rely on the alternate electron acceptors including nitrate (<math>\n <mrow>\n <msubsup>\n <mi>NO</mi>\n <mn>3</mn>\n <mo>−</mo>\n </msubsup>\n </mrow></math>), iron (Fe<sup>3+</sup>), manganese (Mn<sup>4+</sup>), and sulfate (<math>\n <mrow>\n <msubsup>\n <mi>SO</mi>\n <mn>4</mn>\n <mrow>\n <mn>2</mn>\n <mo>−</mo>\n </mrow>\n </msubsup>\n </mrow></math>). These alternate electron acceptors are exploited sequentially based on their energetic favorability and produce electron donors including reduced bioavailable forms of carbon, nitrogen (<math>\n <mrow>\n <msubsup>\n <mi>NH</mi>\n <mn>4</mn>\n <mo>+</mo>\n </msubsup>\n </mrow></math>), iron (Fe<sup>2+</sup>), and sulfur (H<sub>2</sub>S) that may diffuse into overlying oxygenated water (McClure et al. <span>2021</span>). For example, nitrate and ammonium become bioavailable in locations where oxygen conditions fluctuate and allow temporal and spatial coupling of the anaerobic and aerobic metabolisms involved in nitrogen cycling (Davidson et al. <span>2000</span>). In addition, oxygen-depleted water in the sediments can result in high phosphorus concentrations because ferric iron reduction (Fe<sup>3+</sup> to Fe<sup>2+</sup>) coupled with slightly acidic pH conditions (particularly at night when photosynthesis ceases) releases bioavailable forms of phosphorus (Gotoh and Patrick <span>1974</span>). Importantly, diurnal heating of sediments in the littoral zone can drive water movement within surficial sediments, creating convection of pore water that may further increase availability of bioavailable nutrients to periphyton (Axler and Reuter <span>1996</span>).</p><p>Altogether, the fluctuations in oxygen availability and temperature in littoral zones can increase availability of nutrients released from sediment, where they are efficiently intercepted by periphyton (Carlton and Wetzel <span>1988</span>; Dodds <span>2003</span>) and may not become elevated in the water column (e.g., Rosenberger et al. <span>2008</span>). Complex successional patterns, such as diatomaceous mucilage production can initiate an early, physical barrier between the water column and the sediments (Hoagland et al. <span>1982</span>; Wood et al. <span>2015</span>). As the biofilm undergoes colonization and succession, warm temperatures may accelerate heterotrophic processes, thereby promoting biofilm growth and further impeding exchange between the water column and sediment. Most importantly, nutrients released from the sediment are directly used by periphyton in the mat, and do not reach the water column; concurrently, the biofilm impedes movement of oxygen from the water column into the sediment, promoting anoxia which encourages nutrient release.</p><p>Once a robust biofilm is established, periphyton blooms can promote an environment that is favorable for their further growth—both by stimulating internal nutrient release (Carlton and Wetzel <span>1988</span>) and discouraging grazers (Wood et al. <span>2015</span>). The dead cells accumulating at the sediment surface provide additional substrate for microbial growth and respiration, promoting anoxia that releases additional nutrients from the sediment and organic matter (Fig. 1). The steep nutrient gradient between pore water and the overlying water column fosters more rapid movement of nutrients to attached algae, stimulating further growth. During the day, the top of the photosynthetic mat produces abundant oxygen that can be drawn downward across a relatively steep oxygen gradient toward respiring microbes. High pH associated with high oxygen of the photosynthesizing mat can discourage daytime P release from calcareous sediments (Dodds <span>2003</span>). However, at night when photosynthesis ceases, the mat can become hypoxic or anoxic, allowing further release of nutrients from the sediment into the overlying periphyton mat. This strong diel contrast in oxygen and the associated pH not only affects nutrient dynamics but also creates a hostile environment for some grazers (Wood et al. <span>2015</span>) that already may have difficulty consuming the types of filamentous algae that sometimes dominate these blooms (Rosenberger et al. <span>2008</span>). Thus, the periphyton mat is positioned to both take advantage of internal nutrient release and potentially to benefit from reduced grazing pressure, altogether generating favorable conditions for its continued growth.</p><p>Where does this self-reinforcing cycle end? Some have noted that winter temperature and light reductions, as well as wave exposure, can dislodge periphyton mats when dead matter at the base of the mat reduces its stability (Vadeboncoeur et al. <span>2021</span>). In mountain lakes, a spring freshet after a high-snowpack winter can effectively scour material and flush it downstream, such that spring periphyton may have to become established on relatively bare substrate. If not, it is possible that a periphyton mat may persist through the winter or, under snow and ice, die back while still attached, encouraging hypoxia.</p><p>We have demonstrated in principle that microbially mediated nutrient release in tandem with the physical constraints of temperature-dependent oxygen saturation may be a ubiquitous mechanism for driving nearshore benthic algal blooms in clear lakes that are warming, but the data necessary to test this hypothesis are lacking. Limnology has focused on offshore water column dynamics, infrequently including nearshore or benthic measurements, much less those across the varied substrates of littoral zones (Vadeboncoeur et al. <span>2021</span>; Atkins et al. <span>2022</span>). Where data exist, we see that clear mountain lakes can exhibit higher temperatures nearshore, relative to offshore water, but temperature and oxygen measurements that include the sediment and the overlying water column are uncommon. Competing processes could drive water column oxygen nearshore in multiple directions (e.g., wind and waves increase it, daytime periphyton photosynthesis increases it, decomposition and still conditions with a thermal bar decrease it) so these data are clearly needed. In nearshore zones, we anticipate the benthos to be much warmer than the overlying water, as it is warmed not only by the overlying water mass but also by the heat of decomposition. Furthermore, the benthos is warmed by radiant heat from the sun that may be particularly strong at high elevation and particularly effective on darker substrate.</p><p>New research is needed at the sediment–water interface of lakes, to examine temperature and oxygen dynamics, as well as richer studies of nutrient flux and microbial dynamics. We have highlighted processes that may have contributed to the observations of periphyton blooms in relatively well-protected mountain lakes, but also recognize that these dynamics should be relevant for low-elevation lakes where increased periphyton blooms have been reported in recent decades. Warming temperatures, increasing levels of hypoxia, and associated changes in redox conditions may be occurring without detection in lakes where studies focus on the water column rather than the sediment–water interface. Such effects of warming on redox conditions are undoubtedly altering biogeochemistry at global scales, with implications not only for nutrient dynamics but also carbon cycles and pollutant release (Jane et al. <span>2021</span>).</p>","PeriodicalId":18128,"journal":{"name":"Limnology and Oceanography Letters","volume":"9 1","pages":"1-6"},"PeriodicalIF":5.1000,"publicationDate":"2023-10-31","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1002/lol2.10357","citationCount":"0","resultStr":"{\"title\":\"Warming-induced changes in benthic redox as a potential driver of increasing benthic algal blooms in high-elevation lakes\",\"authors\":\"Stephanie E. Hampton, Jill S. Baron, Robert Ladwig, Ryan P. McClure, Michael F. Meyer, Isabella A. Oleksy, Anna Shampain\",\"doi\":\"10.1002/lol2.10357\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p>Blooms of algae attached to the bottom of nearshore environments have increasingly been observed in clear, nutrient-poor lakes worldwide (Vadeboncoeur et al. <span>2021</span>). The phenomenon is puzzling for at least two reasons. First, such nearshore benthic blooms appear to be common across heterogeneous contexts—from the world's largest lake in Siberia to small mountain lakes in the Rocky Mountains of North America. Second, the blooms are enigmatic because they have been observed in the near absence of direct human disturbances, such as those in well-protected, remote mountain lakes. A recent review (Vadeboncoeur et al. <span>2021</span>) highlights the diversity of mechanisms through which these nearshore benthic blooms may be triggered—such as nutrient loading through atmospheric deposition or human activity within the watershed, changes in hydrodynamics associated with climate change, and food web alterations that reduce grazing pressure. Yet none of these potential drivers are common across the lakes where nearshore benthic algal blooms have been reported. Here, we suggest that a previously unappreciated driver is common across all lakes that experience warming associated with climate change—elevated nearshore temperatures that reduce oxygen and promote nutrient release from sediments under the biofilm. In principle, this phenomenon should be particularly notable in high-elevation lakes because water at higher elevation holds less oxygen at saturation, high-elevation lakes are warming more quickly than their lowland counterparts, and they can receive additional warming from relatively high solar radiation (Fig. 1). We briefly review the evidence in support of this concept, recognizing that the data necessary to rigorously test it do not yet exist; however, such data collection is feasible.</p><p>Algal taxa in oligotrophic systems are well adapted to naturally high-light and nutrient-poor conditions (Cantonati and Lowe <span>2014</span>). Because nutrients at the sediment–water interface can be rapidly cycled, nutrient concentrations in the water column may not be reliable indicators of nutrient availability for benthic primary producers (Vadeboncoeur and Power <span>2017</span>). Hot spots or hot moments of elevated nutrient concentrations in littoral regions favor green algae, leading to thick filamentous mats (also called filamentous algal blooms; Oleksy et al. <span>2021</span>; Vadeboncoeur et al. <span>2021</span>). These periphyton blooms appear to be an emergent response to changing environmental conditions in oligotrophic systems, yet the ultimate drivers behind their increased occurrence are debated and often obfuscated by confounding processes. For example, in Lake Baikal, increasing evidence has pointed to highly localized wastewater inputs leading to increased <i>Ulothrix</i> spp. and <i>Spirogyra</i> spp. abundance (Timoshkin et al. <span>2018</span>; Meyer et al. <span>2022</span>). Similarly, Lake Tahoe has experienced increased periphyton biomass likely due to a mix of wastewater-associated eutrophication, groundwater leachate, and complex physical and hydrological processes (Naranjo et al. <span>2019</span>; Atkins et al. <span>2022</span>). Although observations of increased periphyton blooms in lakes with adjacent human population can draw links between human activity and in-lake periphyton responses (e.g., Rosenberger et al. <span>2008</span>), these blooms also appear to be increasing in remote, high-elevation systems that lack permanent human settlements (Oleksy et al. <span>2020</span>).</p><p>High-elevation lakes located in mountainous regions are frequently characterized by low-nutrient concentrations, long periods of ice cover, high solar and UV radiation, as well as heterogeneous landscapes associated with complex topography (Moser et al. <span>2019</span>). Despite their remote locations, historically oligotrophic high-elevation lakes have experienced primary productivity increases that sometimes have been attributed to global change, including atmospheric deposition of reactive nitrogen and phosphorus, species introductions, and climate change (Ballantyne et al. <span>2011</span>; Saros et al. <span>2011</span>; Catalan et al. <span>2013</span>; Brahney et al. <span>2014</span>; Sadro et al. <span>2018</span>; Vadeboncoeur et al. <span>2021</span>). Most of this literature describes changes to water column productivity. However, numerous studies have shown that the frequently neglected benthic habitats of oligotrophic lakes, including those at high elevations, dominate ecosystem processes of primary production and respiration (reviewed by Vadeboncoeur and Power <span>2017</span>). Changes in benthic algal assemblages previously may have been overlooked. For instance, within lakes in the southern Rocky Mountains of North America, sediment records indicate that nearshore algal assemblages have changed appreciably in recent decades, with 200–300% increases in periphytic green algal species (Oleksy et al. <span>2020</span>).</p><p>Although sources and magnitude of nutrient inputs vary greatly across lakes worldwide, a common factor is warming associated with climate change (O'Reilly et al. <span>2015</span>), which can promote oxygen depletion (Jane et al. <span>2021</span>). Mountainous regions are warming faster than adjacent low-lying areas (Pepin et al. <span>2022</span>), and warm water holds less oxygen than cold water. In a high-elevation lake, relative to its lower elevation counterparts, the water column holds lower oxygen at saturation due to the inverse relationship between elevation (atmospheric pressure) and the oxygen capacity of water (Fig. 1). Hutchinson (<span>1957</span>) briefly alluded to the potential for this elevation–oxygen relationship to alter benthic ecology. The importance of not only oxygen content but also partial pressure is increasingly recognized as affecting biodiversity patterns among aquatic invertebrates (e.g., Verberk et al. <span>2011</span>). Our mass balance calculation using a vertical 1D reaction–diffusion model suggests that elevation effects on oxygen in a warming mountain lake could account for sediment anoxia occurring days to weeks earlier than would occur in a warming lake at sea level (Supporting Information). High solar radiation in mountain lakes can contribute to additional warming, and nearshore temperatures can increase markedly relative to the offshore. In some cases, as nearshore temperatures warm, a thermal bar may develop, reducing mixing and allowing littoral water temperatures to become even warmer (Vadeboncoeur et al. <span>2021</span>). Warming sediments can quickly stimulate heterotrophic microbial processes that consume oxygen (Bergström et al. <span>2010</span>). Convective cooling in the littoral zones can further limit horizontal exchange of oxygen and nutrients with offshore waters (Doda et al. <span>2023</span>). These combined factors could explain why mountain lake nearshore blooms have been so noticeable in the absence of other clear drivers.</p><p>Warming littoral zone temperatures can decrease oxygen available for aerobic metabolism, creating an environment more favorable to anaerobic metabolism just a few millimeters into the sediments (Fig. 1, Schlesinger and Bernhardt <span>2013</span>), particularly at night when photosynthesis does not produce oxygen. Microbial consumption of oxygen in sediments frequently exceeds the oxygen supply through diffusion (Fenchel and Finlay <span>2008</span>). Without oxygen, microbes cannot use oxidative phosphorylation to convert organic matter into CO<sub>2</sub>, and instead must rely on the alternate electron acceptors including nitrate (<math>\\n <mrow>\\n <msubsup>\\n <mi>NO</mi>\\n <mn>3</mn>\\n <mo>−</mo>\\n </msubsup>\\n </mrow></math>), iron (Fe<sup>3+</sup>), manganese (Mn<sup>4+</sup>), and sulfate (<math>\\n <mrow>\\n <msubsup>\\n <mi>SO</mi>\\n <mn>4</mn>\\n <mrow>\\n <mn>2</mn>\\n <mo>−</mo>\\n </mrow>\\n </msubsup>\\n </mrow></math>). These alternate electron acceptors are exploited sequentially based on their energetic favorability and produce electron donors including reduced bioavailable forms of carbon, nitrogen (<math>\\n <mrow>\\n <msubsup>\\n <mi>NH</mi>\\n <mn>4</mn>\\n <mo>+</mo>\\n </msubsup>\\n </mrow></math>), iron (Fe<sup>2+</sup>), and sulfur (H<sub>2</sub>S) that may diffuse into overlying oxygenated water (McClure et al. <span>2021</span>). For example, nitrate and ammonium become bioavailable in locations where oxygen conditions fluctuate and allow temporal and spatial coupling of the anaerobic and aerobic metabolisms involved in nitrogen cycling (Davidson et al. <span>2000</span>). In addition, oxygen-depleted water in the sediments can result in high phosphorus concentrations because ferric iron reduction (Fe<sup>3+</sup> to Fe<sup>2+</sup>) coupled with slightly acidic pH conditions (particularly at night when photosynthesis ceases) releases bioavailable forms of phosphorus (Gotoh and Patrick <span>1974</span>). Importantly, diurnal heating of sediments in the littoral zone can drive water movement within surficial sediments, creating convection of pore water that may further increase availability of bioavailable nutrients to periphyton (Axler and Reuter <span>1996</span>).</p><p>Altogether, the fluctuations in oxygen availability and temperature in littoral zones can increase availability of nutrients released from sediment, where they are efficiently intercepted by periphyton (Carlton and Wetzel <span>1988</span>; Dodds <span>2003</span>) and may not become elevated in the water column (e.g., Rosenberger et al. <span>2008</span>). Complex successional patterns, such as diatomaceous mucilage production can initiate an early, physical barrier between the water column and the sediments (Hoagland et al. <span>1982</span>; Wood et al. <span>2015</span>). As the biofilm undergoes colonization and succession, warm temperatures may accelerate heterotrophic processes, thereby promoting biofilm growth and further impeding exchange between the water column and sediment. Most importantly, nutrients released from the sediment are directly used by periphyton in the mat, and do not reach the water column; concurrently, the biofilm impedes movement of oxygen from the water column into the sediment, promoting anoxia which encourages nutrient release.</p><p>Once a robust biofilm is established, periphyton blooms can promote an environment that is favorable for their further growth—both by stimulating internal nutrient release (Carlton and Wetzel <span>1988</span>) and discouraging grazers (Wood et al. <span>2015</span>). The dead cells accumulating at the sediment surface provide additional substrate for microbial growth and respiration, promoting anoxia that releases additional nutrients from the sediment and organic matter (Fig. 1). The steep nutrient gradient between pore water and the overlying water column fosters more rapid movement of nutrients to attached algae, stimulating further growth. During the day, the top of the photosynthetic mat produces abundant oxygen that can be drawn downward across a relatively steep oxygen gradient toward respiring microbes. High pH associated with high oxygen of the photosynthesizing mat can discourage daytime P release from calcareous sediments (Dodds <span>2003</span>). However, at night when photosynthesis ceases, the mat can become hypoxic or anoxic, allowing further release of nutrients from the sediment into the overlying periphyton mat. This strong diel contrast in oxygen and the associated pH not only affects nutrient dynamics but also creates a hostile environment for some grazers (Wood et al. <span>2015</span>) that already may have difficulty consuming the types of filamentous algae that sometimes dominate these blooms (Rosenberger et al. <span>2008</span>). Thus, the periphyton mat is positioned to both take advantage of internal nutrient release and potentially to benefit from reduced grazing pressure, altogether generating favorable conditions for its continued growth.</p><p>Where does this self-reinforcing cycle end? Some have noted that winter temperature and light reductions, as well as wave exposure, can dislodge periphyton mats when dead matter at the base of the mat reduces its stability (Vadeboncoeur et al. <span>2021</span>). In mountain lakes, a spring freshet after a high-snowpack winter can effectively scour material and flush it downstream, such that spring periphyton may have to become established on relatively bare substrate. If not, it is possible that a periphyton mat may persist through the winter or, under snow and ice, die back while still attached, encouraging hypoxia.</p><p>We have demonstrated in principle that microbially mediated nutrient release in tandem with the physical constraints of temperature-dependent oxygen saturation may be a ubiquitous mechanism for driving nearshore benthic algal blooms in clear lakes that are warming, but the data necessary to test this hypothesis are lacking. Limnology has focused on offshore water column dynamics, infrequently including nearshore or benthic measurements, much less those across the varied substrates of littoral zones (Vadeboncoeur et al. <span>2021</span>; Atkins et al. <span>2022</span>). Where data exist, we see that clear mountain lakes can exhibit higher temperatures nearshore, relative to offshore water, but temperature and oxygen measurements that include the sediment and the overlying water column are uncommon. Competing processes could drive water column oxygen nearshore in multiple directions (e.g., wind and waves increase it, daytime periphyton photosynthesis increases it, decomposition and still conditions with a thermal bar decrease it) so these data are clearly needed. In nearshore zones, we anticipate the benthos to be much warmer than the overlying water, as it is warmed not only by the overlying water mass but also by the heat of decomposition. Furthermore, the benthos is warmed by radiant heat from the sun that may be particularly strong at high elevation and particularly effective on darker substrate.</p><p>New research is needed at the sediment–water interface of lakes, to examine temperature and oxygen dynamics, as well as richer studies of nutrient flux and microbial dynamics. We have highlighted processes that may have contributed to the observations of periphyton blooms in relatively well-protected mountain lakes, but also recognize that these dynamics should be relevant for low-elevation lakes where increased periphyton blooms have been reported in recent decades. Warming temperatures, increasing levels of hypoxia, and associated changes in redox conditions may be occurring without detection in lakes where studies focus on the water column rather than the sediment–water interface. Such effects of warming on redox conditions are undoubtedly altering biogeochemistry at global scales, with implications not only for nutrient dynamics but also carbon cycles and pollutant release (Jane et al. <span>2021</span>).</p>\",\"PeriodicalId\":18128,\"journal\":{\"name\":\"Limnology and Oceanography Letters\",\"volume\":\"9 1\",\"pages\":\"1-6\"},\"PeriodicalIF\":5.1000,\"publicationDate\":\"2023-10-31\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://onlinelibrary.wiley.com/doi/epdf/10.1002/lol2.10357\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"Limnology and Oceanography Letters\",\"FirstCategoryId\":\"93\",\"ListUrlMain\":\"https://onlinelibrary.wiley.com/doi/10.1002/lol2.10357\",\"RegionNum\":2,\"RegionCategory\":\"地球科学\",\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"Q1\",\"JCRName\":\"LIMNOLOGY\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"Limnology and Oceanography Letters","FirstCategoryId":"93","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1002/lol2.10357","RegionNum":2,"RegionCategory":"地球科学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"LIMNOLOGY","Score":null,"Total":0}
Warming-induced changes in benthic redox as a potential driver of increasing benthic algal blooms in high-elevation lakes
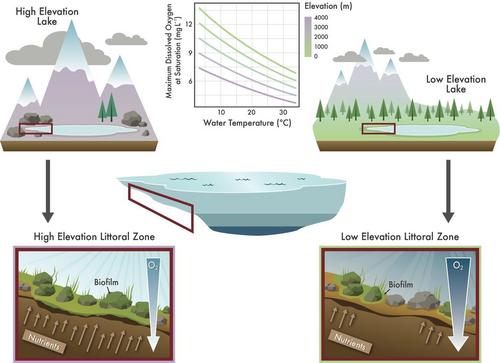
Blooms of algae attached to the bottom of nearshore environments have increasingly been observed in clear, nutrient-poor lakes worldwide (Vadeboncoeur et al. 2021). The phenomenon is puzzling for at least two reasons. First, such nearshore benthic blooms appear to be common across heterogeneous contexts—from the world's largest lake in Siberia to small mountain lakes in the Rocky Mountains of North America. Second, the blooms are enigmatic because they have been observed in the near absence of direct human disturbances, such as those in well-protected, remote mountain lakes. A recent review (Vadeboncoeur et al. 2021) highlights the diversity of mechanisms through which these nearshore benthic blooms may be triggered—such as nutrient loading through atmospheric deposition or human activity within the watershed, changes in hydrodynamics associated with climate change, and food web alterations that reduce grazing pressure. Yet none of these potential drivers are common across the lakes where nearshore benthic algal blooms have been reported. Here, we suggest that a previously unappreciated driver is common across all lakes that experience warming associated with climate change—elevated nearshore temperatures that reduce oxygen and promote nutrient release from sediments under the biofilm. In principle, this phenomenon should be particularly notable in high-elevation lakes because water at higher elevation holds less oxygen at saturation, high-elevation lakes are warming more quickly than their lowland counterparts, and they can receive additional warming from relatively high solar radiation (Fig. 1). We briefly review the evidence in support of this concept, recognizing that the data necessary to rigorously test it do not yet exist; however, such data collection is feasible.
Algal taxa in oligotrophic systems are well adapted to naturally high-light and nutrient-poor conditions (Cantonati and Lowe 2014). Because nutrients at the sediment–water interface can be rapidly cycled, nutrient concentrations in the water column may not be reliable indicators of nutrient availability for benthic primary producers (Vadeboncoeur and Power 2017). Hot spots or hot moments of elevated nutrient concentrations in littoral regions favor green algae, leading to thick filamentous mats (also called filamentous algal blooms; Oleksy et al. 2021; Vadeboncoeur et al. 2021). These periphyton blooms appear to be an emergent response to changing environmental conditions in oligotrophic systems, yet the ultimate drivers behind their increased occurrence are debated and often obfuscated by confounding processes. For example, in Lake Baikal, increasing evidence has pointed to highly localized wastewater inputs leading to increased Ulothrix spp. and Spirogyra spp. abundance (Timoshkin et al. 2018; Meyer et al. 2022). Similarly, Lake Tahoe has experienced increased periphyton biomass likely due to a mix of wastewater-associated eutrophication, groundwater leachate, and complex physical and hydrological processes (Naranjo et al. 2019; Atkins et al. 2022). Although observations of increased periphyton blooms in lakes with adjacent human population can draw links between human activity and in-lake periphyton responses (e.g., Rosenberger et al. 2008), these blooms also appear to be increasing in remote, high-elevation systems that lack permanent human settlements (Oleksy et al. 2020).
High-elevation lakes located in mountainous regions are frequently characterized by low-nutrient concentrations, long periods of ice cover, high solar and UV radiation, as well as heterogeneous landscapes associated with complex topography (Moser et al. 2019). Despite their remote locations, historically oligotrophic high-elevation lakes have experienced primary productivity increases that sometimes have been attributed to global change, including atmospheric deposition of reactive nitrogen and phosphorus, species introductions, and climate change (Ballantyne et al. 2011; Saros et al. 2011; Catalan et al. 2013; Brahney et al. 2014; Sadro et al. 2018; Vadeboncoeur et al. 2021). Most of this literature describes changes to water column productivity. However, numerous studies have shown that the frequently neglected benthic habitats of oligotrophic lakes, including those at high elevations, dominate ecosystem processes of primary production and respiration (reviewed by Vadeboncoeur and Power 2017). Changes in benthic algal assemblages previously may have been overlooked. For instance, within lakes in the southern Rocky Mountains of North America, sediment records indicate that nearshore algal assemblages have changed appreciably in recent decades, with 200–300% increases in periphytic green algal species (Oleksy et al. 2020).
Although sources and magnitude of nutrient inputs vary greatly across lakes worldwide, a common factor is warming associated with climate change (O'Reilly et al. 2015), which can promote oxygen depletion (Jane et al. 2021). Mountainous regions are warming faster than adjacent low-lying areas (Pepin et al. 2022), and warm water holds less oxygen than cold water. In a high-elevation lake, relative to its lower elevation counterparts, the water column holds lower oxygen at saturation due to the inverse relationship between elevation (atmospheric pressure) and the oxygen capacity of water (Fig. 1). Hutchinson (1957) briefly alluded to the potential for this elevation–oxygen relationship to alter benthic ecology. The importance of not only oxygen content but also partial pressure is increasingly recognized as affecting biodiversity patterns among aquatic invertebrates (e.g., Verberk et al. 2011). Our mass balance calculation using a vertical 1D reaction–diffusion model suggests that elevation effects on oxygen in a warming mountain lake could account for sediment anoxia occurring days to weeks earlier than would occur in a warming lake at sea level (Supporting Information). High solar radiation in mountain lakes can contribute to additional warming, and nearshore temperatures can increase markedly relative to the offshore. In some cases, as nearshore temperatures warm, a thermal bar may develop, reducing mixing and allowing littoral water temperatures to become even warmer (Vadeboncoeur et al. 2021). Warming sediments can quickly stimulate heterotrophic microbial processes that consume oxygen (Bergström et al. 2010). Convective cooling in the littoral zones can further limit horizontal exchange of oxygen and nutrients with offshore waters (Doda et al. 2023). These combined factors could explain why mountain lake nearshore blooms have been so noticeable in the absence of other clear drivers.
Warming littoral zone temperatures can decrease oxygen available for aerobic metabolism, creating an environment more favorable to anaerobic metabolism just a few millimeters into the sediments (Fig. 1, Schlesinger and Bernhardt 2013), particularly at night when photosynthesis does not produce oxygen. Microbial consumption of oxygen in sediments frequently exceeds the oxygen supply through diffusion (Fenchel and Finlay 2008). Without oxygen, microbes cannot use oxidative phosphorylation to convert organic matter into CO2, and instead must rely on the alternate electron acceptors including nitrate (), iron (Fe3+), manganese (Mn4+), and sulfate (). These alternate electron acceptors are exploited sequentially based on their energetic favorability and produce electron donors including reduced bioavailable forms of carbon, nitrogen (), iron (Fe2+), and sulfur (H2S) that may diffuse into overlying oxygenated water (McClure et al. 2021). For example, nitrate and ammonium become bioavailable in locations where oxygen conditions fluctuate and allow temporal and spatial coupling of the anaerobic and aerobic metabolisms involved in nitrogen cycling (Davidson et al. 2000). In addition, oxygen-depleted water in the sediments can result in high phosphorus concentrations because ferric iron reduction (Fe3+ to Fe2+) coupled with slightly acidic pH conditions (particularly at night when photosynthesis ceases) releases bioavailable forms of phosphorus (Gotoh and Patrick 1974). Importantly, diurnal heating of sediments in the littoral zone can drive water movement within surficial sediments, creating convection of pore water that may further increase availability of bioavailable nutrients to periphyton (Axler and Reuter 1996).
Altogether, the fluctuations in oxygen availability and temperature in littoral zones can increase availability of nutrients released from sediment, where they are efficiently intercepted by periphyton (Carlton and Wetzel 1988; Dodds 2003) and may not become elevated in the water column (e.g., Rosenberger et al. 2008). Complex successional patterns, such as diatomaceous mucilage production can initiate an early, physical barrier between the water column and the sediments (Hoagland et al. 1982; Wood et al. 2015). As the biofilm undergoes colonization and succession, warm temperatures may accelerate heterotrophic processes, thereby promoting biofilm growth and further impeding exchange between the water column and sediment. Most importantly, nutrients released from the sediment are directly used by periphyton in the mat, and do not reach the water column; concurrently, the biofilm impedes movement of oxygen from the water column into the sediment, promoting anoxia which encourages nutrient release.
Once a robust biofilm is established, periphyton blooms can promote an environment that is favorable for their further growth—both by stimulating internal nutrient release (Carlton and Wetzel 1988) and discouraging grazers (Wood et al. 2015). The dead cells accumulating at the sediment surface provide additional substrate for microbial growth and respiration, promoting anoxia that releases additional nutrients from the sediment and organic matter (Fig. 1). The steep nutrient gradient between pore water and the overlying water column fosters more rapid movement of nutrients to attached algae, stimulating further growth. During the day, the top of the photosynthetic mat produces abundant oxygen that can be drawn downward across a relatively steep oxygen gradient toward respiring microbes. High pH associated with high oxygen of the photosynthesizing mat can discourage daytime P release from calcareous sediments (Dodds 2003). However, at night when photosynthesis ceases, the mat can become hypoxic or anoxic, allowing further release of nutrients from the sediment into the overlying periphyton mat. This strong diel contrast in oxygen and the associated pH not only affects nutrient dynamics but also creates a hostile environment for some grazers (Wood et al. 2015) that already may have difficulty consuming the types of filamentous algae that sometimes dominate these blooms (Rosenberger et al. 2008). Thus, the periphyton mat is positioned to both take advantage of internal nutrient release and potentially to benefit from reduced grazing pressure, altogether generating favorable conditions for its continued growth.
Where does this self-reinforcing cycle end? Some have noted that winter temperature and light reductions, as well as wave exposure, can dislodge periphyton mats when dead matter at the base of the mat reduces its stability (Vadeboncoeur et al. 2021). In mountain lakes, a spring freshet after a high-snowpack winter can effectively scour material and flush it downstream, such that spring periphyton may have to become established on relatively bare substrate. If not, it is possible that a periphyton mat may persist through the winter or, under snow and ice, die back while still attached, encouraging hypoxia.
We have demonstrated in principle that microbially mediated nutrient release in tandem with the physical constraints of temperature-dependent oxygen saturation may be a ubiquitous mechanism for driving nearshore benthic algal blooms in clear lakes that are warming, but the data necessary to test this hypothesis are lacking. Limnology has focused on offshore water column dynamics, infrequently including nearshore or benthic measurements, much less those across the varied substrates of littoral zones (Vadeboncoeur et al. 2021; Atkins et al. 2022). Where data exist, we see that clear mountain lakes can exhibit higher temperatures nearshore, relative to offshore water, but temperature and oxygen measurements that include the sediment and the overlying water column are uncommon. Competing processes could drive water column oxygen nearshore in multiple directions (e.g., wind and waves increase it, daytime periphyton photosynthesis increases it, decomposition and still conditions with a thermal bar decrease it) so these data are clearly needed. In nearshore zones, we anticipate the benthos to be much warmer than the overlying water, as it is warmed not only by the overlying water mass but also by the heat of decomposition. Furthermore, the benthos is warmed by radiant heat from the sun that may be particularly strong at high elevation and particularly effective on darker substrate.
New research is needed at the sediment–water interface of lakes, to examine temperature and oxygen dynamics, as well as richer studies of nutrient flux and microbial dynamics. We have highlighted processes that may have contributed to the observations of periphyton blooms in relatively well-protected mountain lakes, but also recognize that these dynamics should be relevant for low-elevation lakes where increased periphyton blooms have been reported in recent decades. Warming temperatures, increasing levels of hypoxia, and associated changes in redox conditions may be occurring without detection in lakes where studies focus on the water column rather than the sediment–water interface. Such effects of warming on redox conditions are undoubtedly altering biogeochemistry at global scales, with implications not only for nutrient dynamics but also carbon cycles and pollutant release (Jane et al. 2021).
期刊介绍:
Limnology and Oceanography Letters (LO-Letters) serves as a platform for communicating the latest innovative and trend-setting research in the aquatic sciences. Manuscripts submitted to LO-Letters are expected to present high-impact, cutting-edge results, discoveries, or conceptual developments across all areas of limnology and oceanography, including their integration. Selection criteria for manuscripts include their broad relevance to the field, strong empirical and conceptual foundations, succinct and elegant conclusions, and potential to advance knowledge in aquatic sciences.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


