{"title":"根系需要好好修剪以吸收水分吗?","authors":"Yann Boursiac, Fabrice Bauget","doi":"10.1111/nph.19336","DOIUrl":null,"url":null,"abstract":"<p>The roots of vascular plants have evolved tubular outgrowths of their epidermal cells called root hairs. These extensions show large variations between species, accessions, and environmental conditions, in both size and density (Rongsawat <i>et al</i>., <span>2021</span>). They originate from an interplay between epidermal cells and underlying tissues, some of the molecular players acting in their development being well known (Cui <i>et al</i>., <span>2018</span>). Three main functions have been identified so far for these structures: anchorage, biotic interactions, and nutrition (Rongsawat <i>et al</i>., <span>2021</span>). For this last function, it has been shown that root hair development is tuned by nutrient deficiencies. It has also been shown that uptake capacities and the expression of some membrane transporters are specific to root hairs. Therefore, it is rather intuitive to think that, as for other nutrients, root hairs will also be beneficial for water uptake. However, this is not yet clear: experts in this field have generated equivocal experimental and modeling evidences about the beneficial role of root hairs for water uptake (Cai & Ahmed, <span>2022</span>). In an article published in this issue of <i>New Phytologist</i>, Duddek <i>et al</i>. (<span>2023</span>, 2484–2497) propose a workflow using an image-based modeling approach to further pursue work on this topic.</p><p>In their introduction, Duddek <i>et al</i>. clearly and concisely present the current knowledge on the role of root hairs in water uptake. In particular, they refer to experimental results which show, both in laboratory and field studies, as well as in identical or different species, either significant or nonsignificant effects on plant water relations (note that no detrimental effect has been observed). For instance, comparison of water uptake between root hair defective mutants and wild types of barley in water stress condition showed no difference (Dodd & Diatloff, <span>2016</span>), while another study on the same genotypes suggested that root hairs allow higher transpiration rates to be sustained in drying soil (Carminati <i>et al</i>., <span>2017</span>). Indeed, neither the species nor the conditions could clearly explain these different results. This question has also been addressed with a modeling approach. The authors note that 3D image-based modeling of water flow in the soil–root system has been conducted for more than two decades. However, most of the models addressed the difference in hydraulic properties between the bulk soil and the vicinity of the root (the rhizosphere) indirectly. For example, by modifying transport parameters, such as changing the radial conductivity of the root according to the soil water content (Couvreur <i>et al</i>., <span>2014</span>). None of these models has, thus far, been able to take into account the geometrical complexity of root–soil contact because of the poor resolution of images of this zone. With the advances of X-ray CT microtomography, a nondestructive imaging technique able to visualize the internal structure of an object at high resolution from a stack of images, the rhizosphere structure may now be incorporated in models. While previous image-based models focused on the impact of root hairs on nutrient uptake (see, e.g. Keyes <i>et al</i>., <span>2013</span>), or carbon diffusion (Schnepf <i>et al</i>., <span>2022</span>), the novelty of the work presented by Duddek <i>et al</i>. is to use high-resolution (0.65 μm × 0.65 μm per pixel) image-based modeling to study water uptake. This provided an accurate segmentation of the root hairs within the soil micropores, allowing the authors to study the effect of root hairs, and their shrinkage at low soil water potential, on water uptake.</p><p>A 3D reconstruction of six segments of 1.4 mm of maize (<i>Zea mays</i> L.) roots embedded in soil was obtained from synchrotron imaging. The Richards equation, describing the flow of water in unsaturated porous media, was solved on each sample in steady state, and with proper boundary conditions. The parametrization of the soil and root properties was based on previous works (Zarebanadkouki <i>et al</i>., <span>2019</span>; Vetterlein <i>et al</i>., <span>2021</span>), with the assumption that root hairs had the same hydraulic conductivity as cortical cells. Impact of root hairs on water uptake was evaluated by numerically removing them from the reconstructed root segment, with the main benefit being enabling the assessment of the geometrical aspect of their presence/absence while avoiding possible compensation mechanisms that could occur in root hair defective mutants. A significant increase in the water uptake of the root segments in dry soil conditions was found for the hairy case: up to 24% at <i>h</i><sub>in</sub> = −1.56 MPa. This increase would mainly be due to an increase in root–soil contact. Their simulations also showed that the water potential gradient in the vicinity of the root would be lower in the hairy case, leading to a more homogeneous water uptake. The root hair shrinkage effect was evaluated through scenarios in which the root cortex and hair hydraulic conductivity was dependent on the soil water potential. As expected, shrinkage strongly reduced root hair impact on water uptake (Fig. 1).</p><p>Although this work, according to assumptions and parametrization choices, clearly demonstrates the efficiency of root hairs, the authors do not forget to point out some limitations of their approach. For instance, the model was not intended to simulate water uptake at the whole root scale. Indeed, the computational cost at this resolution would be too high. Consequently, at the present scale of a few mm<sup>3</sup>, phenomena like hydrotropism or hydropatterning cannot be considered, neither other soil aspects, such as heterogeneity in porosity. The authors also mention a possible limitation in their current simulations, related to the hydraulic conductivity of root hairs being the same as that of cortical cells. Unfortunately, root hair hydraulic conductivity is not an easy property to measure. The choice here was made based on the literature (Jones <i>et al</i>., <span>1983</span>), but their model will be ready to handle more hydraulic configurations as soon as they become available. Duddek <i>et al</i>. also note the coupling of their model of water flow with other models at the cell scale, such as MECHA (Couvreur <i>et al</i>., <span>2018</span>), would provide the opportunity to distinguish hydraulic properties according to the cell type. Overall, this work carries the idea that modeling approaches are needed, since root hairs are not accessible for functional assays when grown in soil, and alternative growth conditions may alter their properties (Nestler <i>et al</i>., <span>2016</span>).</p><p>Finally, and as is often the case when asking questions in biological sciences, to the question whether root hairs are beneficial for water uptake, the answer of the authors is not ‘yes’ or ‘no’ but rather ‘it depends’. First, they were able to highlight the subtleties required to understand the function of root hairs within root hydraulic architecture and soil. Second, their model provides a way to predict, for a given set of soil and root conductivities, a window of soil conditions in which root hairs can be beneficial.</p><p>Ultimately, the work by Duddek <i>et al</i>. raises an unfortunate paradox for root hairs: they are able to increase root–soil contact, which is critical under low soil water content, but at the same time they will shrink under those conditions, therefore interrupting that very same contact. However, this may not be a paradox. On top of, or instead of, a quantitative role in water uptake, it has been suggested that root hairs could serve as a surveillance system for the soil water status (Kwasniewski <i>et al</i>., <span>2016</span>). Root hairs could therefore play a qualitative role in plant water relations, in a similar way as a transistor, by releasing systemic signaling processes and adaptive responses when they shrink because of low soil water potential (Fig. 1). Future studies will surely shed light on these aspects.</p>","PeriodicalId":214,"journal":{"name":"New Phytologist","volume":"240 6","pages":"2173-2175"},"PeriodicalIF":8.1000,"publicationDate":"2023-10-16","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://nph.onlinelibrary.wiley.com/doi/epdf/10.1111/nph.19336","citationCount":"0","resultStr":"{\"title\":\"Do roots need a good haircut for water uptake?\",\"authors\":\"Yann Boursiac, Fabrice Bauget\",\"doi\":\"10.1111/nph.19336\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p>The roots of vascular plants have evolved tubular outgrowths of their epidermal cells called root hairs. These extensions show large variations between species, accessions, and environmental conditions, in both size and density (Rongsawat <i>et al</i>., <span>2021</span>). They originate from an interplay between epidermal cells and underlying tissues, some of the molecular players acting in their development being well known (Cui <i>et al</i>., <span>2018</span>). Three main functions have been identified so far for these structures: anchorage, biotic interactions, and nutrition (Rongsawat <i>et al</i>., <span>2021</span>). For this last function, it has been shown that root hair development is tuned by nutrient deficiencies. It has also been shown that uptake capacities and the expression of some membrane transporters are specific to root hairs. Therefore, it is rather intuitive to think that, as for other nutrients, root hairs will also be beneficial for water uptake. However, this is not yet clear: experts in this field have generated equivocal experimental and modeling evidences about the beneficial role of root hairs for water uptake (Cai & Ahmed, <span>2022</span>). In an article published in this issue of <i>New Phytologist</i>, Duddek <i>et al</i>. (<span>2023</span>, 2484–2497) propose a workflow using an image-based modeling approach to further pursue work on this topic.</p><p>In their introduction, Duddek <i>et al</i>. clearly and concisely present the current knowledge on the role of root hairs in water uptake. In particular, they refer to experimental results which show, both in laboratory and field studies, as well as in identical or different species, either significant or nonsignificant effects on plant water relations (note that no detrimental effect has been observed). For instance, comparison of water uptake between root hair defective mutants and wild types of barley in water stress condition showed no difference (Dodd & Diatloff, <span>2016</span>), while another study on the same genotypes suggested that root hairs allow higher transpiration rates to be sustained in drying soil (Carminati <i>et al</i>., <span>2017</span>). Indeed, neither the species nor the conditions could clearly explain these different results. This question has also been addressed with a modeling approach. The authors note that 3D image-based modeling of water flow in the soil–root system has been conducted for more than two decades. However, most of the models addressed the difference in hydraulic properties between the bulk soil and the vicinity of the root (the rhizosphere) indirectly. For example, by modifying transport parameters, such as changing the radial conductivity of the root according to the soil water content (Couvreur <i>et al</i>., <span>2014</span>). None of these models has, thus far, been able to take into account the geometrical complexity of root–soil contact because of the poor resolution of images of this zone. With the advances of X-ray CT microtomography, a nondestructive imaging technique able to visualize the internal structure of an object at high resolution from a stack of images, the rhizosphere structure may now be incorporated in models. While previous image-based models focused on the impact of root hairs on nutrient uptake (see, e.g. Keyes <i>et al</i>., <span>2013</span>), or carbon diffusion (Schnepf <i>et al</i>., <span>2022</span>), the novelty of the work presented by Duddek <i>et al</i>. is to use high-resolution (0.65 μm × 0.65 μm per pixel) image-based modeling to study water uptake. This provided an accurate segmentation of the root hairs within the soil micropores, allowing the authors to study the effect of root hairs, and their shrinkage at low soil water potential, on water uptake.</p><p>A 3D reconstruction of six segments of 1.4 mm of maize (<i>Zea mays</i> L.) roots embedded in soil was obtained from synchrotron imaging. The Richards equation, describing the flow of water in unsaturated porous media, was solved on each sample in steady state, and with proper boundary conditions. The parametrization of the soil and root properties was based on previous works (Zarebanadkouki <i>et al</i>., <span>2019</span>; Vetterlein <i>et al</i>., <span>2021</span>), with the assumption that root hairs had the same hydraulic conductivity as cortical cells. Impact of root hairs on water uptake was evaluated by numerically removing them from the reconstructed root segment, with the main benefit being enabling the assessment of the geometrical aspect of their presence/absence while avoiding possible compensation mechanisms that could occur in root hair defective mutants. A significant increase in the water uptake of the root segments in dry soil conditions was found for the hairy case: up to 24% at <i>h</i><sub>in</sub> = −1.56 MPa. This increase would mainly be due to an increase in root–soil contact. Their simulations also showed that the water potential gradient in the vicinity of the root would be lower in the hairy case, leading to a more homogeneous water uptake. The root hair shrinkage effect was evaluated through scenarios in which the root cortex and hair hydraulic conductivity was dependent on the soil water potential. As expected, shrinkage strongly reduced root hair impact on water uptake (Fig. 1).</p><p>Although this work, according to assumptions and parametrization choices, clearly demonstrates the efficiency of root hairs, the authors do not forget to point out some limitations of their approach. For instance, the model was not intended to simulate water uptake at the whole root scale. Indeed, the computational cost at this resolution would be too high. Consequently, at the present scale of a few mm<sup>3</sup>, phenomena like hydrotropism or hydropatterning cannot be considered, neither other soil aspects, such as heterogeneity in porosity. The authors also mention a possible limitation in their current simulations, related to the hydraulic conductivity of root hairs being the same as that of cortical cells. Unfortunately, root hair hydraulic conductivity is not an easy property to measure. The choice here was made based on the literature (Jones <i>et al</i>., <span>1983</span>), but their model will be ready to handle more hydraulic configurations as soon as they become available. Duddek <i>et al</i>. also note the coupling of their model of water flow with other models at the cell scale, such as MECHA (Couvreur <i>et al</i>., <span>2018</span>), would provide the opportunity to distinguish hydraulic properties according to the cell type. Overall, this work carries the idea that modeling approaches are needed, since root hairs are not accessible for functional assays when grown in soil, and alternative growth conditions may alter their properties (Nestler <i>et al</i>., <span>2016</span>).</p><p>Finally, and as is often the case when asking questions in biological sciences, to the question whether root hairs are beneficial for water uptake, the answer of the authors is not ‘yes’ or ‘no’ but rather ‘it depends’. First, they were able to highlight the subtleties required to understand the function of root hairs within root hydraulic architecture and soil. Second, their model provides a way to predict, for a given set of soil and root conductivities, a window of soil conditions in which root hairs can be beneficial.</p><p>Ultimately, the work by Duddek <i>et al</i>. raises an unfortunate paradox for root hairs: they are able to increase root–soil contact, which is critical under low soil water content, but at the same time they will shrink under those conditions, therefore interrupting that very same contact. However, this may not be a paradox. On top of, or instead of, a quantitative role in water uptake, it has been suggested that root hairs could serve as a surveillance system for the soil water status (Kwasniewski <i>et al</i>., <span>2016</span>). Root hairs could therefore play a qualitative role in plant water relations, in a similar way as a transistor, by releasing systemic signaling processes and adaptive responses when they shrink because of low soil water potential (Fig. 1). Future studies will surely shed light on these aspects.</p>\",\"PeriodicalId\":214,\"journal\":{\"name\":\"New Phytologist\",\"volume\":\"240 6\",\"pages\":\"2173-2175\"},\"PeriodicalIF\":8.1000,\"publicationDate\":\"2023-10-16\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://nph.onlinelibrary.wiley.com/doi/epdf/10.1111/nph.19336\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"New Phytologist\",\"FirstCategoryId\":\"99\",\"ListUrlMain\":\"https://onlinelibrary.wiley.com/doi/10.1111/nph.19336\",\"RegionNum\":1,\"RegionCategory\":\"生物学\",\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"Q1\",\"JCRName\":\"PLANT SCIENCES\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"New Phytologist","FirstCategoryId":"99","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1111/nph.19336","RegionNum":1,"RegionCategory":"生物学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"PLANT SCIENCES","Score":null,"Total":0}
引用次数: 0
摘要
维管植物的根已经进化出管状的表皮细胞,称为根毛。这些扩展在物种、添加物和环境条件之间的大小和密度都有很大差异(Rongsawat et al., 2021)。它们起源于表皮细胞和底层组织之间的相互作用,其中一些在其发育过程中起作用的分子是众所周知的(Cui et al., 2018)。到目前为止,已经确定了这些结构的三个主要功能:锚定、生物相互作用和营养(Rongsawat et al., 2021)。对于这最后一个功能,已经证明根毛的发育是由营养缺乏调节的。研究还表明,吸收能力和一些膜转运蛋白的表达是根毛所特有的。因此,我们很直观地认为,与其他营养物质一样,根毛也有利于水分的吸收。然而,这一点尚不清楚:该领域的专家已经得出了关于根毛对水分吸收有益作用的模棱两可的实验和模型证据(Cai &艾哈迈德,2022)。在本期《新植物学家》上发表的一篇文章中,Duddek等人(2023,2484 - 2497)提出了一种使用基于图像的建模方法的工作流程,以进一步研究这一主题。在他们的介绍中,Duddek等人清晰而简洁地介绍了目前关于根毛在水分吸收中的作用的知识。特别是,它们提到的实验结果表明,在实验室和实地研究中,以及在相同或不同的物种中,对植物的水分关系有显著或不显著的影响(注意,没有观察到有害影响)。例如,在水分胁迫条件下,根毛缺陷突变体和野生型大麦的吸水量没有差异(Dodd &Diatloff, 2016),而另一项关于相同基因型的研究表明,根毛可以在干燥的土壤中维持更高的蒸腾速率(Carminati等人,2017)。事实上,无论是物种还是环境都不能清楚地解释这些不同的结果。这个问题也通过建模方法得到了解决。作者指出,基于三维图像的土壤根系水流模型已经进行了20多年。然而,大多数模型间接地解决了散装土壤和根部附近(根际)之间的水力特性差异。例如,通过改变输送参数,如根据土壤含水量改变根系的径向导电性(Couvreur et al., 2014)。由于该区域的图像分辨率较差,迄今为止,这些模型都无法考虑到根与土壤接触的几何复杂性。随着x射线CT微断层扫描技术的进步,一种非破坏性成像技术能够从一堆图像中以高分辨率可视化物体的内部结构,现在可以将根际结构纳入模型。先前基于图像的模型关注的是根毛对养分吸收的影响(例如Keyes等人,2013)或碳扩散(Schnepf等人,2022),Duddek等人提出的工作的新颖之处在于使用高分辨率(0.65 μm × 0.65 μm /像素)基于图像的建模来研究水分吸收。这提供了土壤微孔内根毛的精确分割,使作者能够研究根毛及其在低土壤水势下的收缩对水分吸收的影响。利用同步加速器成像技术对埋于土壤中的6段1.4 mm玉米根系进行了三维重建。描述水在非饱和多孔介质中流动的理查兹方程在稳定状态下和适当的边界条件下对每个样品进行求解。土壤和根系特性的参数化基于之前的工作(Zarebanadkouki等人,2019;Vetterlein et al., 2021),假设根毛与皮质细胞具有相同的水力传导性。通过从重建的根段中数值去除根毛对吸水的影响,评估了根毛对吸水的影响,其主要好处是能够评估其存在/不存在的几何方面,同时避免了可能发生在根毛缺陷突变体中的补偿机制。在干燥的土壤条件下,有毛的情况下,根系的吸水性显著增加:在hin = - 1.56 MPa时,吸水性高达24%。这种增加主要是由于根与土壤接触的增加。他们的模拟还表明,在毛茸茸的情况下,根部附近的水势梯度会更低,从而导致更均匀的水分吸收。通过根皮质和根毛导水率依赖于土壤水势的情景来评价根毛收缩效应。 正如预期的那样,收缩大大降低了根毛对水分吸收的影响(图1)。尽管这项工作,根据假设和参数化选择,清楚地证明了根毛的效率,但作者没有忘记指出他们的方法的一些局限性。例如,该模型并不打算模拟整个根系尺度上的水分吸收。事实上,这种分辨率下的计算成本太高了。因此,在目前的几毫米尺度下,不能考虑亲水或水型化等现象,也不能考虑土壤的其他方面,如孔隙度的非均质性。作者还提到了他们目前的模拟中可能存在的一个限制,即根毛的水力传导性与皮质细胞的相同。不幸的是,根毛的水力导电性不是一个容易测量的特性。这里的选择是基于文献(Jones et al., 1983),但他们的模型将准备处理更多的液压配置,一旦他们成为可用的。Duddek等人还注意到,他们的水流模型与细胞尺度上的其他模型(如MECHA)的耦合(Couvreur等人,2018)将提供根据细胞类型区分水力特性的机会。总的来说,这项工作提出了建模方法是必要的,因为根毛在土壤中生长时无法进行功能分析,而其他生长条件可能会改变其特性(Nestler等人,2016)。最后,就像在生物科学中提出问题时经常出现的情况一样,对于根毛是否有利于水分吸收的问题,作者的回答不是“是”或“否”,而是“视情况而定”。首先,他们能够强调理解根毛在根水力结构和土壤中的功能所需的微妙之处。其次,他们的模型提供了一种方法来预测,对于给定的土壤和根的电导率,一个土壤条件的窗口,在这个窗口中,根毛可能是有益的。最终,Duddek等人的研究为根毛提出了一个不幸的悖论:它们能够增加根与土壤的接触,这在土壤含水量低的情况下是至关重要的,但与此同时,它们会在这些条件下收缩,从而中断了同样的接触。然而,这可能不是一个悖论。除了在水分吸收中的定量作用之外,也有人认为根毛可以作为土壤水分状况的监测系统(Kwasniewski et al., 2016)。因此,根毛可以在植物水分关系中发挥定性作用,以类似于晶体管的方式,通过释放系统信号过程和自适应反应,当它们因土壤水势低而收缩时(图1)。未来的研究肯定会阐明这些方面。
The roots of vascular plants have evolved tubular outgrowths of their epidermal cells called root hairs. These extensions show large variations between species, accessions, and environmental conditions, in both size and density (Rongsawat et al., 2021). They originate from an interplay between epidermal cells and underlying tissues, some of the molecular players acting in their development being well known (Cui et al., 2018). Three main functions have been identified so far for these structures: anchorage, biotic interactions, and nutrition (Rongsawat et al., 2021). For this last function, it has been shown that root hair development is tuned by nutrient deficiencies. It has also been shown that uptake capacities and the expression of some membrane transporters are specific to root hairs. Therefore, it is rather intuitive to think that, as for other nutrients, root hairs will also be beneficial for water uptake. However, this is not yet clear: experts in this field have generated equivocal experimental and modeling evidences about the beneficial role of root hairs for water uptake (Cai & Ahmed, 2022). In an article published in this issue of New Phytologist, Duddek et al. (2023, 2484–2497) propose a workflow using an image-based modeling approach to further pursue work on this topic.
In their introduction, Duddek et al. clearly and concisely present the current knowledge on the role of root hairs in water uptake. In particular, they refer to experimental results which show, both in laboratory and field studies, as well as in identical or different species, either significant or nonsignificant effects on plant water relations (note that no detrimental effect has been observed). For instance, comparison of water uptake between root hair defective mutants and wild types of barley in water stress condition showed no difference (Dodd & Diatloff, 2016), while another study on the same genotypes suggested that root hairs allow higher transpiration rates to be sustained in drying soil (Carminati et al., 2017). Indeed, neither the species nor the conditions could clearly explain these different results. This question has also been addressed with a modeling approach. The authors note that 3D image-based modeling of water flow in the soil–root system has been conducted for more than two decades. However, most of the models addressed the difference in hydraulic properties between the bulk soil and the vicinity of the root (the rhizosphere) indirectly. For example, by modifying transport parameters, such as changing the radial conductivity of the root according to the soil water content (Couvreur et al., 2014). None of these models has, thus far, been able to take into account the geometrical complexity of root–soil contact because of the poor resolution of images of this zone. With the advances of X-ray CT microtomography, a nondestructive imaging technique able to visualize the internal structure of an object at high resolution from a stack of images, the rhizosphere structure may now be incorporated in models. While previous image-based models focused on the impact of root hairs on nutrient uptake (see, e.g. Keyes et al., 2013), or carbon diffusion (Schnepf et al., 2022), the novelty of the work presented by Duddek et al. is to use high-resolution (0.65 μm × 0.65 μm per pixel) image-based modeling to study water uptake. This provided an accurate segmentation of the root hairs within the soil micropores, allowing the authors to study the effect of root hairs, and their shrinkage at low soil water potential, on water uptake.
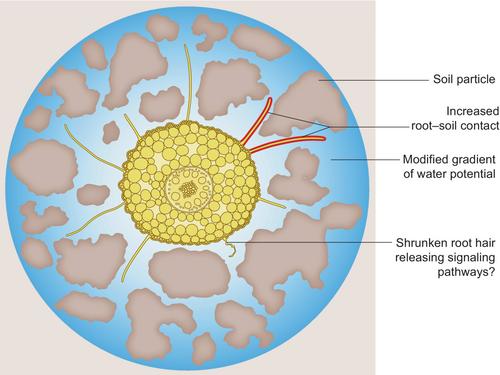
A 3D reconstruction of six segments of 1.4 mm of maize (Zea mays L.) roots embedded in soil was obtained from synchrotron imaging. The Richards equation, describing the flow of water in unsaturated porous media, was solved on each sample in steady state, and with proper boundary conditions. The parametrization of the soil and root properties was based on previous works (Zarebanadkouki et al., 2019; Vetterlein et al., 2021), with the assumption that root hairs had the same hydraulic conductivity as cortical cells. Impact of root hairs on water uptake was evaluated by numerically removing them from the reconstructed root segment, with the main benefit being enabling the assessment of the geometrical aspect of their presence/absence while avoiding possible compensation mechanisms that could occur in root hair defective mutants. A significant increase in the water uptake of the root segments in dry soil conditions was found for the hairy case: up to 24% at hin = −1.56 MPa. This increase would mainly be due to an increase in root–soil contact. Their simulations also showed that the water potential gradient in the vicinity of the root would be lower in the hairy case, leading to a more homogeneous water uptake. The root hair shrinkage effect was evaluated through scenarios in which the root cortex and hair hydraulic conductivity was dependent on the soil water potential. As expected, shrinkage strongly reduced root hair impact on water uptake (Fig. 1).
Although this work, according to assumptions and parametrization choices, clearly demonstrates the efficiency of root hairs, the authors do not forget to point out some limitations of their approach. For instance, the model was not intended to simulate water uptake at the whole root scale. Indeed, the computational cost at this resolution would be too high. Consequently, at the present scale of a few mm3, phenomena like hydrotropism or hydropatterning cannot be considered, neither other soil aspects, such as heterogeneity in porosity. The authors also mention a possible limitation in their current simulations, related to the hydraulic conductivity of root hairs being the same as that of cortical cells. Unfortunately, root hair hydraulic conductivity is not an easy property to measure. The choice here was made based on the literature (Jones et al., 1983), but their model will be ready to handle more hydraulic configurations as soon as they become available. Duddek et al. also note the coupling of their model of water flow with other models at the cell scale, such as MECHA (Couvreur et al., 2018), would provide the opportunity to distinguish hydraulic properties according to the cell type. Overall, this work carries the idea that modeling approaches are needed, since root hairs are not accessible for functional assays when grown in soil, and alternative growth conditions may alter their properties (Nestler et al., 2016).
Finally, and as is often the case when asking questions in biological sciences, to the question whether root hairs are beneficial for water uptake, the answer of the authors is not ‘yes’ or ‘no’ but rather ‘it depends’. First, they were able to highlight the subtleties required to understand the function of root hairs within root hydraulic architecture and soil. Second, their model provides a way to predict, for a given set of soil and root conductivities, a window of soil conditions in which root hairs can be beneficial.
Ultimately, the work by Duddek et al. raises an unfortunate paradox for root hairs: they are able to increase root–soil contact, which is critical under low soil water content, but at the same time they will shrink under those conditions, therefore interrupting that very same contact. However, this may not be a paradox. On top of, or instead of, a quantitative role in water uptake, it has been suggested that root hairs could serve as a surveillance system for the soil water status (Kwasniewski et al., 2016). Root hairs could therefore play a qualitative role in plant water relations, in a similar way as a transistor, by releasing systemic signaling processes and adaptive responses when they shrink because of low soil water potential (Fig. 1). Future studies will surely shed light on these aspects.
期刊介绍:
New Phytologist is an international electronic journal published 24 times a year. It is owned by the New Phytologist Foundation, a non-profit-making charitable organization dedicated to promoting plant science. The journal publishes excellent, novel, rigorous, and timely research and scholarship in plant science and its applications. The articles cover topics in five sections: Physiology & Development, Environment, Interaction, Evolution, and Transformative Plant Biotechnology. These sections encompass intracellular processes, global environmental change, and encourage cross-disciplinary approaches. The journal recognizes the use of techniques from molecular and cell biology, functional genomics, modeling, and system-based approaches in plant science. Abstracting and Indexing Information for New Phytologist includes Academic Search, AgBiotech News & Information, Agroforestry Abstracts, Biochemistry & Biophysics Citation Index, Botanical Pesticides, CAB Abstracts®, Environment Index, Global Health, and Plant Breeding Abstracts, and others.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


