Rose McNelly, Ángel Vergara-Cruces, David Lea-Smith, David Seung, Michael Webster
{"title":"探索质体生物学和生物技术的潜力:质体预览会,诺里奇,2022年9月1日至2日。","authors":"Rose McNelly, Ángel Vergara-Cruces, David Lea-Smith, David Seung, Michael Webster","doi":"10.1111/nph.19296","DOIUrl":null,"url":null,"abstract":"<p>Plastids are a family of organelles that likely originate from the endosymbiosis of cyanobacteria (Howe <i>et al</i>., <span>2008</span>) and exhibit diverse morphologies and biochemical capabilities (Fig. 1). The chloroplast is the most well-studied plastid type and performs photosynthesis in plants and algae and is thus responsible for most of the food production on the planet. However, there are other important biochemical functions performed by plastids, including the synthesis and storage of some carbohydrates, pigment production, fatty acid synthesis and nitrogen and sulphur assimilation. Plants have specialised plastid types that carry out some of these functions, such as starch-storing amyloplasts and pigment-producing chromoplasts (Fig. 1).</p><p>The annual Plastid Preview Meeting took place on 1–2 September 2022 in Norwich (UK), jointly hosted by the John Innes Centre and the University of East Anglia. Since the 1970s, Plastid Preview has showcased cutting-edge research from PhD students and early career postdoctoral scientists investigating fundamental and applied plastid biology. Research presented throughout the years has not only addressed questions in photosynthesis but also in diverse areas such as chlorophyll biosynthesis, starch metabolism, carbon fixation, plastid gene expression and plastid transformation. In the 2022 meeting, it was apparent that significant progress is being made in addressing long-standing questions in these areas with the aid of technological developments, such as machine learning, gene editing and cryo-EM approaches to study protein structure, combined with the ability to explore previously untapped biological diversity with the rapid expansion of genome sequences.</p><p>Three major domains of plastid research were covered: the first concerned ‘understanding the plastid’, probing the fundamental mechanisms that govern plastid biology. The second concerned ‘optimising the plastid’ and focussed on processes that could increase productivity and resilience in cyanobacteria, algae and plants. The third concerned ‘utilising the plastid’, specifically the use of plastid-containing organisms in biotechnology to produce high-value proteins and metabolites. Here, we highlight some of the diverse and exciting areas of work encompassing these three domains that were covered at the meeting.</p><p>Research into fundamental plastid processes, such as biogenesis, gene expression, protein import and division, is required to understand how photosynthesis and metabolism occur and are integrated within the organelle.</p><p>Protein import is a prime example of a process that influences many other plastid functions and was an area of active discussion at the meeting. The majority of the plastid proteome is encoded in the nucleus, and proteins are imported into the organelle through recognition of plastid-localisation signals, translocation through the plastid membranes and delivery to the appropriate location within the plastid. This process heavily influences the plastid proteome and thus plastid function, and recent exciting work implicated ubiquitination in this process (Ling & Jarvis, <span>2015</span>). Ling <i>et al</i>. (<span>2012</span>) discovered that the cytosolic ubiquitin-mediated protein degradation pathway regulates components of the protein import machinery. More recently, ubiquitin-mediated protein degradation has also been observed for a broad range of chloroplast proteins in <i>Arabidopsis</i>, including those involved in photosynthesis, oxidative stress and other metabolic pathways (Sun <i>et al</i>., <span>2022</span>). This opens new and exciting questions about the role of ubiquitination in the regulation of these processes, as well as its relevance to a broader range of species and plastid types, including crops. We also discussed the exciting links that were emerging between fundamental plastid processes, such as between protein import and plastid division (Fang <i>et al</i>., <span>2022</span>).</p><p>While protein import is an example of how the nucleus can influence plastid function, the plastid possesses reciprocal mechanisms to signal its status back to the nucleus, known as retrograde signalling. Many aspects of this communication are not fully understood, but several important components have been uncovered, such as tetrapyrroles and reactive oxygen species (Chan <i>et al</i>., <span>2016</span>). Many of these components accumulate due to a defect in photosynthesis, allowing the nucleus to sense the status of the chloroplast. Since photosynthesis is sensitive to perturbations in environmental conditions, this makes the chloroplast an ideal sentinel for general cell status. As such, retrograde signalling is involved in the response to both abiotic and biotic stresses (Chan <i>et al</i>., <span>2016</span>; Littlejohn <i>et al</i>., <span>2021</span>). An example of emerging research discussed at the meeting is how pathogen infection can affect nuclear expression of genes encoding plastid-localised proteins that influence resistance (Corredor-Moreno <i>et al</i>., <span>2022</span>). Uncovering how pathogen effectors hijack plastid metabolism during infection and the broader roles of the plastid in biotic interactions could lead to novel approaches to protect plants from pathogens.</p><p>Photosynthesis is a central role of chloroplasts and is the main source of energy in cyanobacteria. The inefficiencies of RuBisCo have been described in detail (Flamholz <i>et al</i>., <span>2019</span>). It can be partly attributed to the slow evolution of the enzyme (Bouvier <i>et al</i>., <span>2022</span>) and the requirement to balance carboxylation rates with CO<sub>2</sub>/O<sub>2</sub> specificity. The meeting highlighted exciting strategies to overcome these and increase RuBisCo activity.</p><p>The first strategy was based on harnessing pre-existing diversity in RuBisCo. There is large variation in RuBisCo kinetics between phototrophs (Savir <i>et al</i>., <span>2010</span>) that could potentially be exploited to replace native enzymes with more efficient versions, if localised CO<sub>2</sub> concentrations could be increased. Extensive modelling shows that replacing the wheat RuBisCo with that from a C<sub>4</sub> species could significantly increase the efficiency of carbon uptake (Iqbal <i>et al</i>., <span>2021</span>), and experimentally implementing these predictions may validate this route to increasing photosynthetic efficiency. Furthermore, there are many species where RuBisCo kinetics have not been reported, particularly in nonmodel species. As machine learning becomes increasingly prevalent, it may be employed to predict RuBisCo sequences with the potential to improve photosynthesis.</p><p>The second strategy is to improve RuBisCo efficiency via carbon concentrating mechanisms (CCMs). CCMs can be biophysical, like algal pyrenoids and cyanobacterial carboxysomes, or biochemical, as in C<sub>4</sub> plants. Recent work has focussed on developing a mechanistic understanding on how these CCMs form and function. One breakthrough was the discovery of the <i>Chlamydomonas reinhardtii</i> protein EPYC1, which physically links RuBisCo to the pyrenoid (Mackinder <i>et al</i>., <span>2016</span>). This not only enhanced our understanding of pyrenoid assembly but also opened the possibility of engineering pyrenoids into plants (Adler <i>et al</i>., <span>2022</span>). Similar work to transfer carboxysomes of cyanobacteria into plants may follow, benefitting from recent insights into the structure of the α-carboxysome (Ni <i>et al</i>., <span>2022</span>).</p><p>Beyond photosynthesis in chloroplasts, there are many metabolic pathways that could be manipulated to increase crop quality, from carotenoid storage in chromoplasts to fatty acid synthesis in elaioplasts (Fig. 1). Specifically, exciting progress has been made on starch synthesis in amyloplasts of wheat, which produce two distinct types of starch granules, large A-type and small B-type granules. The discovery that the protein MRC regulates the formation of B-type granules can be exploited to introduce desirable changes in starch granule size properties for the food and milling industries (Chen <i>et al</i>., <span>2022</span>). Further understanding of these processes may come from exploiting intra- and interspecies diversity and will drive improvements in crop nutritional and functional quality in a similar way to improvements in photosynthesis.</p><p>Biotechnological approaches to using the plastid as a biofactory have been a key theme of recent Plastid Preview meetings. The plastid provides a distinct environment from the rest of the cell for producing valuable proteins and metabolites. Much attention has fallen on cyanobacteria and algae, especially newly discovered fast-growing species like <i>Synechococcus</i> sp. PCC 11901 (Włodarczyk <i>et al</i>., <span>2020</span>), for which large volumes of cells can be cultivated with ease and in a smaller space compared with plants. There are numerous examples of <i>C. reinhardtii</i> chloroplast engineering to produce vaccine subunits, antibacterial compounds and antibodies (Dyo & Purton, <span>2018</span>).</p><p>A primary focus of this meeting was to explore new techniques and approaches to improve cyanobacterial, algal and plant biotechnology. Regulators of chloroplast transgenes are of great interest, to allow robust control and easy on/off switching of expression. Riboswitches, for instance the thiamine pyrophosphate riboswitch, are a potential solution (Mehrshahi <i>et al</i>., <span>2020</span>), and the discovery of further novel switches and using these in tandem can expand our existing toolkit. Another area highlighted was targeting heterologous proteins to distinct plastid subcompartments, like the chloroplast thylakoid lumen. This would expand the range of proteins that can be produced to those that are not stable in the stroma (Chin-Fatt & Menassa, <span>2021</span>).</p><p>To ensure that such exciting biotechnological approaches can be realised, a key priority is to expand our toolkit of plastid transformation technologies. This includes improved synthetic biology tools, for example, MoClo cloning systems that are compatible across a range of phototrophs (Vasudevan <i>et al</i>., <span>2019</span>). We expect that progress will come from developing novel Cas enzymes, beyond Cas9, for efficient function in the plastid and adapting base editing technology for efficient plastome editing. An additional challenge when engineering organisms with multiple plastids per cell is achieving homoplasmy, and exciting new techniques are emerging to overcome this (Okuzaki <i>et al</i>., <span>2020</span>). These methods could result in the widespread adoption of plastid transformation and utilisation of plastids as biofactories in cyanobacteria, algae and plants.</p><p>The Plastid Preview Meeting remains guided by its fundamental goal of shining a light on plastid function, both in photosynthesis and beyond. This meeting highlighted diversity in the plastid field – from the variety of plastid types studied, to intra- and interspecies differences in mechanisms of plastid function. However, much natural diversity remains unexplored and technologies continue to evolve, particularly through adopting and combining techniques from different fields. Exploiting these will be essential to understand, improve and utilise the plastid further, and we have no doubt that this will be covered at future Plastid Previews.</p><p>RM and ÁV-C contributed equally to this work.</p>","PeriodicalId":214,"journal":{"name":"New Phytologist","volume":"240 6","pages":"2187-2190"},"PeriodicalIF":8.3000,"publicationDate":"2023-10-03","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://nph.onlinelibrary.wiley.com/doi/epdf/10.1111/nph.19296","citationCount":"0","resultStr":"{\"title\":\"Exploring the potential of plastid biology and biotechnology\",\"authors\":\"Rose McNelly, Ángel Vergara-Cruces, David Lea-Smith, David Seung, Michael Webster\",\"doi\":\"10.1111/nph.19296\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p>Plastids are a family of organelles that likely originate from the endosymbiosis of cyanobacteria (Howe <i>et al</i>., <span>2008</span>) and exhibit diverse morphologies and biochemical capabilities (Fig. 1). The chloroplast is the most well-studied plastid type and performs photosynthesis in plants and algae and is thus responsible for most of the food production on the planet. However, there are other important biochemical functions performed by plastids, including the synthesis and storage of some carbohydrates, pigment production, fatty acid synthesis and nitrogen and sulphur assimilation. Plants have specialised plastid types that carry out some of these functions, such as starch-storing amyloplasts and pigment-producing chromoplasts (Fig. 1).</p><p>The annual Plastid Preview Meeting took place on 1–2 September 2022 in Norwich (UK), jointly hosted by the John Innes Centre and the University of East Anglia. Since the 1970s, Plastid Preview has showcased cutting-edge research from PhD students and early career postdoctoral scientists investigating fundamental and applied plastid biology. Research presented throughout the years has not only addressed questions in photosynthesis but also in diverse areas such as chlorophyll biosynthesis, starch metabolism, carbon fixation, plastid gene expression and plastid transformation. In the 2022 meeting, it was apparent that significant progress is being made in addressing long-standing questions in these areas with the aid of technological developments, such as machine learning, gene editing and cryo-EM approaches to study protein structure, combined with the ability to explore previously untapped biological diversity with the rapid expansion of genome sequences.</p><p>Three major domains of plastid research were covered: the first concerned ‘understanding the plastid’, probing the fundamental mechanisms that govern plastid biology. The second concerned ‘optimising the plastid’ and focussed on processes that could increase productivity and resilience in cyanobacteria, algae and plants. The third concerned ‘utilising the plastid’, specifically the use of plastid-containing organisms in biotechnology to produce high-value proteins and metabolites. Here, we highlight some of the diverse and exciting areas of work encompassing these three domains that were covered at the meeting.</p><p>Research into fundamental plastid processes, such as biogenesis, gene expression, protein import and division, is required to understand how photosynthesis and metabolism occur and are integrated within the organelle.</p><p>Protein import is a prime example of a process that influences many other plastid functions and was an area of active discussion at the meeting. The majority of the plastid proteome is encoded in the nucleus, and proteins are imported into the organelle through recognition of plastid-localisation signals, translocation through the plastid membranes and delivery to the appropriate location within the plastid. This process heavily influences the plastid proteome and thus plastid function, and recent exciting work implicated ubiquitination in this process (Ling & Jarvis, <span>2015</span>). Ling <i>et al</i>. (<span>2012</span>) discovered that the cytosolic ubiquitin-mediated protein degradation pathway regulates components of the protein import machinery. More recently, ubiquitin-mediated protein degradation has also been observed for a broad range of chloroplast proteins in <i>Arabidopsis</i>, including those involved in photosynthesis, oxidative stress and other metabolic pathways (Sun <i>et al</i>., <span>2022</span>). This opens new and exciting questions about the role of ubiquitination in the regulation of these processes, as well as its relevance to a broader range of species and plastid types, including crops. We also discussed the exciting links that were emerging between fundamental plastid processes, such as between protein import and plastid division (Fang <i>et al</i>., <span>2022</span>).</p><p>While protein import is an example of how the nucleus can influence plastid function, the plastid possesses reciprocal mechanisms to signal its status back to the nucleus, known as retrograde signalling. Many aspects of this communication are not fully understood, but several important components have been uncovered, such as tetrapyrroles and reactive oxygen species (Chan <i>et al</i>., <span>2016</span>). Many of these components accumulate due to a defect in photosynthesis, allowing the nucleus to sense the status of the chloroplast. Since photosynthesis is sensitive to perturbations in environmental conditions, this makes the chloroplast an ideal sentinel for general cell status. As such, retrograde signalling is involved in the response to both abiotic and biotic stresses (Chan <i>et al</i>., <span>2016</span>; Littlejohn <i>et al</i>., <span>2021</span>). An example of emerging research discussed at the meeting is how pathogen infection can affect nuclear expression of genes encoding plastid-localised proteins that influence resistance (Corredor-Moreno <i>et al</i>., <span>2022</span>). Uncovering how pathogen effectors hijack plastid metabolism during infection and the broader roles of the plastid in biotic interactions could lead to novel approaches to protect plants from pathogens.</p><p>Photosynthesis is a central role of chloroplasts and is the main source of energy in cyanobacteria. The inefficiencies of RuBisCo have been described in detail (Flamholz <i>et al</i>., <span>2019</span>). It can be partly attributed to the slow evolution of the enzyme (Bouvier <i>et al</i>., <span>2022</span>) and the requirement to balance carboxylation rates with CO<sub>2</sub>/O<sub>2</sub> specificity. The meeting highlighted exciting strategies to overcome these and increase RuBisCo activity.</p><p>The first strategy was based on harnessing pre-existing diversity in RuBisCo. There is large variation in RuBisCo kinetics between phototrophs (Savir <i>et al</i>., <span>2010</span>) that could potentially be exploited to replace native enzymes with more efficient versions, if localised CO<sub>2</sub> concentrations could be increased. Extensive modelling shows that replacing the wheat RuBisCo with that from a C<sub>4</sub> species could significantly increase the efficiency of carbon uptake (Iqbal <i>et al</i>., <span>2021</span>), and experimentally implementing these predictions may validate this route to increasing photosynthetic efficiency. Furthermore, there are many species where RuBisCo kinetics have not been reported, particularly in nonmodel species. As machine learning becomes increasingly prevalent, it may be employed to predict RuBisCo sequences with the potential to improve photosynthesis.</p><p>The second strategy is to improve RuBisCo efficiency via carbon concentrating mechanisms (CCMs). CCMs can be biophysical, like algal pyrenoids and cyanobacterial carboxysomes, or biochemical, as in C<sub>4</sub> plants. Recent work has focussed on developing a mechanistic understanding on how these CCMs form and function. One breakthrough was the discovery of the <i>Chlamydomonas reinhardtii</i> protein EPYC1, which physically links RuBisCo to the pyrenoid (Mackinder <i>et al</i>., <span>2016</span>). This not only enhanced our understanding of pyrenoid assembly but also opened the possibility of engineering pyrenoids into plants (Adler <i>et al</i>., <span>2022</span>). Similar work to transfer carboxysomes of cyanobacteria into plants may follow, benefitting from recent insights into the structure of the α-carboxysome (Ni <i>et al</i>., <span>2022</span>).</p><p>Beyond photosynthesis in chloroplasts, there are many metabolic pathways that could be manipulated to increase crop quality, from carotenoid storage in chromoplasts to fatty acid synthesis in elaioplasts (Fig. 1). Specifically, exciting progress has been made on starch synthesis in amyloplasts of wheat, which produce two distinct types of starch granules, large A-type and small B-type granules. The discovery that the protein MRC regulates the formation of B-type granules can be exploited to introduce desirable changes in starch granule size properties for the food and milling industries (Chen <i>et al</i>., <span>2022</span>). Further understanding of these processes may come from exploiting intra- and interspecies diversity and will drive improvements in crop nutritional and functional quality in a similar way to improvements in photosynthesis.</p><p>Biotechnological approaches to using the plastid as a biofactory have been a key theme of recent Plastid Preview meetings. The plastid provides a distinct environment from the rest of the cell for producing valuable proteins and metabolites. Much attention has fallen on cyanobacteria and algae, especially newly discovered fast-growing species like <i>Synechococcus</i> sp. PCC 11901 (Włodarczyk <i>et al</i>., <span>2020</span>), for which large volumes of cells can be cultivated with ease and in a smaller space compared with plants. There are numerous examples of <i>C. reinhardtii</i> chloroplast engineering to produce vaccine subunits, antibacterial compounds and antibodies (Dyo & Purton, <span>2018</span>).</p><p>A primary focus of this meeting was to explore new techniques and approaches to improve cyanobacterial, algal and plant biotechnology. Regulators of chloroplast transgenes are of great interest, to allow robust control and easy on/off switching of expression. Riboswitches, for instance the thiamine pyrophosphate riboswitch, are a potential solution (Mehrshahi <i>et al</i>., <span>2020</span>), and the discovery of further novel switches and using these in tandem can expand our existing toolkit. Another area highlighted was targeting heterologous proteins to distinct plastid subcompartments, like the chloroplast thylakoid lumen. This would expand the range of proteins that can be produced to those that are not stable in the stroma (Chin-Fatt & Menassa, <span>2021</span>).</p><p>To ensure that such exciting biotechnological approaches can be realised, a key priority is to expand our toolkit of plastid transformation technologies. This includes improved synthetic biology tools, for example, MoClo cloning systems that are compatible across a range of phototrophs (Vasudevan <i>et al</i>., <span>2019</span>). We expect that progress will come from developing novel Cas enzymes, beyond Cas9, for efficient function in the plastid and adapting base editing technology for efficient plastome editing. An additional challenge when engineering organisms with multiple plastids per cell is achieving homoplasmy, and exciting new techniques are emerging to overcome this (Okuzaki <i>et al</i>., <span>2020</span>). These methods could result in the widespread adoption of plastid transformation and utilisation of plastids as biofactories in cyanobacteria, algae and plants.</p><p>The Plastid Preview Meeting remains guided by its fundamental goal of shining a light on plastid function, both in photosynthesis and beyond. This meeting highlighted diversity in the plastid field – from the variety of plastid types studied, to intra- and interspecies differences in mechanisms of plastid function. However, much natural diversity remains unexplored and technologies continue to evolve, particularly through adopting and combining techniques from different fields. Exploiting these will be essential to understand, improve and utilise the plastid further, and we have no doubt that this will be covered at future Plastid Previews.</p><p>RM and ÁV-C contributed equally to this work.</p>\",\"PeriodicalId\":214,\"journal\":{\"name\":\"New Phytologist\",\"volume\":\"240 6\",\"pages\":\"2187-2190\"},\"PeriodicalIF\":8.3000,\"publicationDate\":\"2023-10-03\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://nph.onlinelibrary.wiley.com/doi/epdf/10.1111/nph.19296\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"New Phytologist\",\"FirstCategoryId\":\"99\",\"ListUrlMain\":\"https://onlinelibrary.wiley.com/doi/10.1111/nph.19296\",\"RegionNum\":1,\"RegionCategory\":\"生物学\",\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"Q1\",\"JCRName\":\"PLANT SCIENCES\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"New Phytologist","FirstCategoryId":"99","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1111/nph.19296","RegionNum":1,"RegionCategory":"生物学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"PLANT SCIENCES","Score":null,"Total":0}
引用次数: 0
摘要
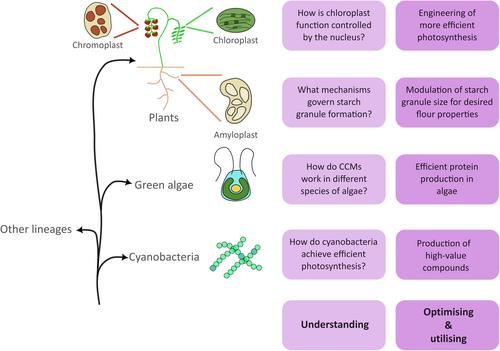
质体是一个细胞器家族,可能起源于蓝藻的内共生(Howe et al., 2008),并表现出多种形态和生化能力(图1)。叶绿体是研究最多的质体类型,在植物和藻类中进行光合作用,因此负责地球上大部分的食物生产。然而,质体还具有其他重要的生化功能,包括一些碳水化合物的合成和储存、色素的产生、脂肪酸的合成以及氮和硫的同化。植物有专门的质体类型来执行这些功能,比如储存淀粉的淀粉质体和产生色素的色质体(图1)。一年一度的质体预演会议于2022年9月1日至2日在诺里奇(英国)举行,由约翰英纳斯中心和东安格利亚大学联合主办。自20世纪70年代以来,质体预览展示了博士生和早期职业博士后科学家研究基础和应用质体生物学的前沿研究。多年来提出的研究不仅涉及光合作用的问题,还涉及叶绿素生物合成、淀粉代谢、碳固定、质体基因表达和质体转化等多个领域。在2022年的会议上,很明显,在技术发展的帮助下,在解决这些领域长期存在的问题方面取得了重大进展,例如机器学习、基因编辑和冷冻电镜研究蛋白质结构的方法,以及随着基因组序列的快速扩展,探索以前未开发的生物多样性的能力。会议涵盖了质体研究的三个主要领域:第一个领域涉及“理解质体”,探索控制质体生物学的基本机制。第二项研究涉及“优化质体”,重点关注可以提高蓝藻、藻类和植物的生产力和复原力的过程。第三个涉及“利用质体”,特别是在生物技术中使用含有质体的生物体来生产高价值的蛋白质和代谢物。在这里,我们将重点介绍会议上涉及的这三个领域的一些多样化和令人兴奋的工作领域。研究质体的基本过程,如生物发生、基因表达、蛋白质输入和分裂,是了解光合作用和代谢如何发生和整合在细胞器内的必要条件。蛋白质输入是影响许多其他质体功能的过程的一个主要例子,也是会议上积极讨论的一个领域。大部分质体蛋白质组在细胞核中编码,蛋白质通过识别质体定位信号,通过质体膜易位并传递到质体内的适当位置,从而进入细胞器。这一过程严重影响质体蛋白质组,从而影响质体功能,最近令人兴奋的研究表明泛素化在这一过程中(Ling &贾维斯,2015)。Ling等人(2012)发现胞质泛素介导的蛋白质降解途径调节蛋白质进口机制的组成部分。最近,在拟南芥中,泛素介导的蛋白质降解也被观察到,包括参与光合作用、氧化应激和其他代谢途径的叶绿体蛋白质(Sun et al., 2022)。这开启了新的和令人兴奋的问题,关于泛素化在这些过程的调节中的作用,以及它与包括作物在内的更广泛的物种和质体类型的相关性。我们还讨论了基本质体过程之间出现的令人兴奋的联系,例如蛋白质输入和质体分裂之间的联系(Fang et al., 2022)。虽然蛋白质输入是细胞核如何影响质体功能的一个例子,但质体具有相互机制,将其状态信号传回细胞核,称为逆行信号。这种交流的许多方面尚未完全了解,但已经发现了几个重要的组成部分,例如四吡啶和活性氧(Chan et al., 2016)。由于光合作用的缺陷,许多这些成分积累起来,使细胞核能够感知叶绿体的状态。由于光合作用对环境条件的扰动很敏感,这使得叶绿体成为一般细胞状态的理想哨兵。因此,逆行信号参与了对非生物和生物胁迫的反应(Chan et al., 2016;Littlejohn et al., 2021)。会议讨论的新兴研究的一个例子是病原体感染如何影响编码影响抗性的质体定位蛋白的基因的核表达(Corredor-Moreno et al., 2022)。 揭示病原体效应物如何在感染过程中劫持质体代谢,以及质体在生物相互作用中的更广泛作用,可能会带来保护植物免受病原体侵害的新方法。光合作用是叶绿体的核心作用,是蓝藻的主要能量来源。RuBisCo的低效率已经被详细描述过(Flamholz et al., 2019)。这可以部分归因于酶的缓慢进化(Bouvier et al., 2022)以及平衡羧化率与CO2/O2特异性的需求。会议强调了克服这些问题和增加RuBisCo活性的令人兴奋的策略。第一种策略是基于利用RuBisCo中已有的多样性。光养生物之间的RuBisCo动力学存在很大差异(Savir等人,2010),如果可以增加局部二氧化碳浓度,可能会利用更有效的版本取代天然酶。广泛的建模表明,用C4物种的RuBisCo取代小麦RuBisCo可以显著提高碳吸收效率(Iqbal等人,2021),并且通过实验实施这些预测可以验证这一途径提高光合效率。此外,还有许多物种的RuBisCo动力学尚未报道,特别是在非模式物种中。随着机器学习变得越来越普遍,它可能被用来预测具有改善光合作用潜力的RuBisCo序列。第二个策略是通过碳浓缩机制(CCMs)提高RuBisCo效率。CCMs可以是生物物理的,如藻类类pyrenoids和蓝藻羧酶体,也可以是生物化学的,如C4植物。最近的工作集中在发展对这些ccm如何形成和功能的机制理解上。其中一个突破是发现了莱茵衣藻蛋白EPYC1,它将RuBisCo与类pyrenoid物理连接起来(Mackinder et al., 2016)。这不仅增强了我们对类芘组装的理解,而且开启了将类芘工程引入植物的可能性(Adler等人,2022)。类似的将蓝藻的羧酸体转移到植物中的工作可能会随之而来,这得益于最近对α-羧酸体结构的见解(Ni et al., 2022)。除了叶绿体的光合作用外,还有许多代谢途径可以通过控制来提高作物质量,从色质体中的类胡萝卜素储存到叶绿体中的脂肪酸合成(图1)。具体来说,小麦淀粉质体的淀粉合成取得了令人兴奋的进展,淀粉质体产生两种不同类型的淀粉颗粒,大a型和小b型颗粒。蛋白质MRC调节b型颗粒形成的发现可以用来为食品和碾磨行业引入淀粉颗粒大小特性的理想变化(Chen et al., 2022)。对这些过程的进一步了解可能来自于利用种内和种间多样性,并将以类似于改善光合作用的方式推动作物营养和功能质量的改善。利用质体作为生物工厂的生物技术方法是最近质体预览会议的一个关键主题。质体为产生有价值的蛋白质和代谢物提供了与细胞其他部分不同的环境。蓝藻和藻类备受关注,尤其是新发现的快速生长物种,如聚藻球菌(Synechococcus sp. PCC 11901) (Włodarczyk et al., 2020),与植物相比,它们可以在更小的空间内轻松培养大量细胞。有许多莱茵哈氏梭菌叶绿体工程产生疫苗亚基、抗菌化合物和抗体的例子(Dyo &Purton, 2018)。本次会议的主要焦点是探索新的技术和方法,以改善蓝藻,藻类和植物生物技术。叶绿体转基因的调节因子是非常有趣的,它允许强大的控制和容易的表达开关。核开关,例如硫胺素焦磷酸核开关,是一种潜在的解决方案(Mehrshahi等人,2020),发现进一步的新型开关并串联使用这些开关可以扩展我们现有的工具包。另一个突出的领域是将异源蛋白靶向到不同的质体亚室,如叶绿体类囊体管腔。这将扩大可以产生的蛋白质的范围,使之成为那些在基质中不稳定的蛋白质(chin - fat;Menassa, 2021)。为了确保这些令人兴奋的生物技术方法能够实现,一个关键的优先事项是扩展我们的质体转化技术工具包。这包括改进的合成生物学工具,例如,在一系列光养生物中兼容的MoClo克隆系统(Vasudevan等人,2019)。 我们期望开发新的Cas酶,超越Cas9,在质体中有效地发挥作用,并采用碱基编辑技术进行高效的质体编辑,从而取得进展。当每个细胞具有多个质体的工程生物体实现同质性时,另一个挑战是令人兴奋的新技术正在出现以克服这一问题(Okuzaki et al., 2020)。这些方法可以导致广泛采用质体转化和利用质体作为生物工厂在蓝藻,藻类和植物。质体预演会议的基本目标是揭示质体在光合作用及其他方面的功能。这次会议强调了质体领域的多样性——从所研究的质体类型的多样性,到质体功能机制的种内和种间差异。然而,许多自然多样性仍未开发,技术继续发展,特别是通过采用和结合来自不同领域的技术。利用这些将是必不可少的理解,改进和利用质体进一步,我们毫不怀疑,这将涵盖在未来的质体预览。RM和ÁV-C对这项工作贡献相同。
Exploring the potential of plastid biology and biotechnology
Plastids are a family of organelles that likely originate from the endosymbiosis of cyanobacteria (Howe et al., 2008) and exhibit diverse morphologies and biochemical capabilities (Fig. 1). The chloroplast is the most well-studied plastid type and performs photosynthesis in plants and algae and is thus responsible for most of the food production on the planet. However, there are other important biochemical functions performed by plastids, including the synthesis and storage of some carbohydrates, pigment production, fatty acid synthesis and nitrogen and sulphur assimilation. Plants have specialised plastid types that carry out some of these functions, such as starch-storing amyloplasts and pigment-producing chromoplasts (Fig. 1).
The annual Plastid Preview Meeting took place on 1–2 September 2022 in Norwich (UK), jointly hosted by the John Innes Centre and the University of East Anglia. Since the 1970s, Plastid Preview has showcased cutting-edge research from PhD students and early career postdoctoral scientists investigating fundamental and applied plastid biology. Research presented throughout the years has not only addressed questions in photosynthesis but also in diverse areas such as chlorophyll biosynthesis, starch metabolism, carbon fixation, plastid gene expression and plastid transformation. In the 2022 meeting, it was apparent that significant progress is being made in addressing long-standing questions in these areas with the aid of technological developments, such as machine learning, gene editing and cryo-EM approaches to study protein structure, combined with the ability to explore previously untapped biological diversity with the rapid expansion of genome sequences.
Three major domains of plastid research were covered: the first concerned ‘understanding the plastid’, probing the fundamental mechanisms that govern plastid biology. The second concerned ‘optimising the plastid’ and focussed on processes that could increase productivity and resilience in cyanobacteria, algae and plants. The third concerned ‘utilising the plastid’, specifically the use of plastid-containing organisms in biotechnology to produce high-value proteins and metabolites. Here, we highlight some of the diverse and exciting areas of work encompassing these three domains that were covered at the meeting.
Research into fundamental plastid processes, such as biogenesis, gene expression, protein import and division, is required to understand how photosynthesis and metabolism occur and are integrated within the organelle.
Protein import is a prime example of a process that influences many other plastid functions and was an area of active discussion at the meeting. The majority of the plastid proteome is encoded in the nucleus, and proteins are imported into the organelle through recognition of plastid-localisation signals, translocation through the plastid membranes and delivery to the appropriate location within the plastid. This process heavily influences the plastid proteome and thus plastid function, and recent exciting work implicated ubiquitination in this process (Ling & Jarvis, 2015). Ling et al. (2012) discovered that the cytosolic ubiquitin-mediated protein degradation pathway regulates components of the protein import machinery. More recently, ubiquitin-mediated protein degradation has also been observed for a broad range of chloroplast proteins in Arabidopsis, including those involved in photosynthesis, oxidative stress and other metabolic pathways (Sun et al., 2022). This opens new and exciting questions about the role of ubiquitination in the regulation of these processes, as well as its relevance to a broader range of species and plastid types, including crops. We also discussed the exciting links that were emerging between fundamental plastid processes, such as between protein import and plastid division (Fang et al., 2022).
While protein import is an example of how the nucleus can influence plastid function, the plastid possesses reciprocal mechanisms to signal its status back to the nucleus, known as retrograde signalling. Many aspects of this communication are not fully understood, but several important components have been uncovered, such as tetrapyrroles and reactive oxygen species (Chan et al., 2016). Many of these components accumulate due to a defect in photosynthesis, allowing the nucleus to sense the status of the chloroplast. Since photosynthesis is sensitive to perturbations in environmental conditions, this makes the chloroplast an ideal sentinel for general cell status. As such, retrograde signalling is involved in the response to both abiotic and biotic stresses (Chan et al., 2016; Littlejohn et al., 2021). An example of emerging research discussed at the meeting is how pathogen infection can affect nuclear expression of genes encoding plastid-localised proteins that influence resistance (Corredor-Moreno et al., 2022). Uncovering how pathogen effectors hijack plastid metabolism during infection and the broader roles of the plastid in biotic interactions could lead to novel approaches to protect plants from pathogens.
Photosynthesis is a central role of chloroplasts and is the main source of energy in cyanobacteria. The inefficiencies of RuBisCo have been described in detail (Flamholz et al., 2019). It can be partly attributed to the slow evolution of the enzyme (Bouvier et al., 2022) and the requirement to balance carboxylation rates with CO2/O2 specificity. The meeting highlighted exciting strategies to overcome these and increase RuBisCo activity.
The first strategy was based on harnessing pre-existing diversity in RuBisCo. There is large variation in RuBisCo kinetics between phototrophs (Savir et al., 2010) that could potentially be exploited to replace native enzymes with more efficient versions, if localised CO2 concentrations could be increased. Extensive modelling shows that replacing the wheat RuBisCo with that from a C4 species could significantly increase the efficiency of carbon uptake (Iqbal et al., 2021), and experimentally implementing these predictions may validate this route to increasing photosynthetic efficiency. Furthermore, there are many species where RuBisCo kinetics have not been reported, particularly in nonmodel species. As machine learning becomes increasingly prevalent, it may be employed to predict RuBisCo sequences with the potential to improve photosynthesis.
The second strategy is to improve RuBisCo efficiency via carbon concentrating mechanisms (CCMs). CCMs can be biophysical, like algal pyrenoids and cyanobacterial carboxysomes, or biochemical, as in C4 plants. Recent work has focussed on developing a mechanistic understanding on how these CCMs form and function. One breakthrough was the discovery of the Chlamydomonas reinhardtii protein EPYC1, which physically links RuBisCo to the pyrenoid (Mackinder et al., 2016). This not only enhanced our understanding of pyrenoid assembly but also opened the possibility of engineering pyrenoids into plants (Adler et al., 2022). Similar work to transfer carboxysomes of cyanobacteria into plants may follow, benefitting from recent insights into the structure of the α-carboxysome (Ni et al., 2022).
Beyond photosynthesis in chloroplasts, there are many metabolic pathways that could be manipulated to increase crop quality, from carotenoid storage in chromoplasts to fatty acid synthesis in elaioplasts (Fig. 1). Specifically, exciting progress has been made on starch synthesis in amyloplasts of wheat, which produce two distinct types of starch granules, large A-type and small B-type granules. The discovery that the protein MRC regulates the formation of B-type granules can be exploited to introduce desirable changes in starch granule size properties for the food and milling industries (Chen et al., 2022). Further understanding of these processes may come from exploiting intra- and interspecies diversity and will drive improvements in crop nutritional and functional quality in a similar way to improvements in photosynthesis.
Biotechnological approaches to using the plastid as a biofactory have been a key theme of recent Plastid Preview meetings. The plastid provides a distinct environment from the rest of the cell for producing valuable proteins and metabolites. Much attention has fallen on cyanobacteria and algae, especially newly discovered fast-growing species like Synechococcus sp. PCC 11901 (Włodarczyk et al., 2020), for which large volumes of cells can be cultivated with ease and in a smaller space compared with plants. There are numerous examples of C. reinhardtii chloroplast engineering to produce vaccine subunits, antibacterial compounds and antibodies (Dyo & Purton, 2018).
A primary focus of this meeting was to explore new techniques and approaches to improve cyanobacterial, algal and plant biotechnology. Regulators of chloroplast transgenes are of great interest, to allow robust control and easy on/off switching of expression. Riboswitches, for instance the thiamine pyrophosphate riboswitch, are a potential solution (Mehrshahi et al., 2020), and the discovery of further novel switches and using these in tandem can expand our existing toolkit. Another area highlighted was targeting heterologous proteins to distinct plastid subcompartments, like the chloroplast thylakoid lumen. This would expand the range of proteins that can be produced to those that are not stable in the stroma (Chin-Fatt & Menassa, 2021).
To ensure that such exciting biotechnological approaches can be realised, a key priority is to expand our toolkit of plastid transformation technologies. This includes improved synthetic biology tools, for example, MoClo cloning systems that are compatible across a range of phototrophs (Vasudevan et al., 2019). We expect that progress will come from developing novel Cas enzymes, beyond Cas9, for efficient function in the plastid and adapting base editing technology for efficient plastome editing. An additional challenge when engineering organisms with multiple plastids per cell is achieving homoplasmy, and exciting new techniques are emerging to overcome this (Okuzaki et al., 2020). These methods could result in the widespread adoption of plastid transformation and utilisation of plastids as biofactories in cyanobacteria, algae and plants.
The Plastid Preview Meeting remains guided by its fundamental goal of shining a light on plastid function, both in photosynthesis and beyond. This meeting highlighted diversity in the plastid field – from the variety of plastid types studied, to intra- and interspecies differences in mechanisms of plastid function. However, much natural diversity remains unexplored and technologies continue to evolve, particularly through adopting and combining techniques from different fields. Exploiting these will be essential to understand, improve and utilise the plastid further, and we have no doubt that this will be covered at future Plastid Previews.
期刊介绍:
New Phytologist is an international electronic journal published 24 times a year. It is owned by the New Phytologist Foundation, a non-profit-making charitable organization dedicated to promoting plant science. The journal publishes excellent, novel, rigorous, and timely research and scholarship in plant science and its applications. The articles cover topics in five sections: Physiology & Development, Environment, Interaction, Evolution, and Transformative Plant Biotechnology. These sections encompass intracellular processes, global environmental change, and encourage cross-disciplinary approaches. The journal recognizes the use of techniques from molecular and cell biology, functional genomics, modeling, and system-based approaches in plant science. Abstracting and Indexing Information for New Phytologist includes Academic Search, AgBiotech News & Information, Agroforestry Abstracts, Biochemistry & Biophysics Citation Index, Botanical Pesticides, CAB Abstracts®, Environment Index, Global Health, and Plant Breeding Abstracts, and others.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


