Art Daniel, Luke St Heaps, Dianne Sylvester, Sara Diaz, Gregory Peters
{"title":"合子后产生的两个嵌合末端反向复制:新端粒可能形成的证据。","authors":"Art Daniel, Luke St Heaps, Dianne Sylvester, Sara Diaz, Gregory Peters","doi":"10.1186/1475-9268-7-1","DOIUrl":null,"url":null,"abstract":"<p><strong>Objective: </strong>To elucidate the structure of terminal inverted duplications and to investigate potential mechanisms of formation in two cases where there was mosaicism with cells of apparently normal karyotype.</p><p><strong>Results: </strong>A karyotype [46,XY,inv dup(4)(p16.3p15.1)/46,XY] performed on blood lymphocytes from a patient referred for developmental delay (case 1) demonstrated a normal karyotype in 60% of cells with a terminal inverted duplication 4p in the remainder. In case 2, referred for multiple fetal anomalies on an ultrasound scan, 33% of amniocyte colonies were karyotypically normal, with a terminal inv dup 10p in the remainder [46,XX,inv dup(10)(p15.3p11)/46,XX]. Duplicated FISH signals for GATA3 and NEBL loci (in case 2), and for the Wolf-Hirschhorn locus (case 1) confirmed the inverted structure of both duplications. In the GTL banded normal cells from both cases, there was a cryptic deletion detected by FISH of one copy of the subtelomere 4p (case 1, probe GS-36P21), and subtelomere 10p (case 2, probe GS-306F7). At pter on both inv dup chromosomes there was no FISH signal present for the specific subtelomere probe. However, a positive pantelomeric probe signal was detected at 4 pter and 10 pter in both the cryptically-deleted chromosomes and the inv dup chromosomes in the respective cell lines of both cases.</p><p><strong>Conclusion: </strong>An inv dup structure was evident for both cases on GTL bands, and confirmed by the various FISH studies. The presence of telomere (TTAGGG repeat) sequences at pter on the inv dup chromosomes (where more proximal chromosome specific subtelomeric probes were negative) was indicated by the pantelomeric probe signals in both cases. We conclude the most likely mechanism of origin in both cases was by sub-telomeric breakage in the zygote at pter, and delayed repair/rearrangement until after one or more subsequent mitotic divisions. In these divisions, at least one breakage-fusion-bridge cycle occurred, to produce inverted duplications. It is proposed then that two differently \"repaired\" daughter cells proliferated in parallel. In one daughter cell line (with an overtly normal karyotype) there was deletion of the subtelomere and presumed repair through capping by a neo-telomere (i.e. \"healing\", as initially proposed by McClintock). This occurred in both cases presented. In the other daughter cell of each case, it is proposed that chromosome stabilization was achieved (after replication) by sister chromatid reunion to form a dicentric, which broke at a subsequent anaphase, to form an inverted duplication (with loss of the reciprocal product, and the other daughter cell line). One inv dup was repaired without an interstitial specific subtelomere (case 1) and one was repaired with a duplicated specific interstitial subtelomere (case 2). After repair TTAGGG repeats were detected by FISH at each respective new pter.</p>","PeriodicalId":84415,"journal":{"name":"Cell & chromosome","volume":"7 ","pages":"1"},"PeriodicalIF":0.0000,"publicationDate":"2008-03-10","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://sci-hub-pdf.com/10.1186/1475-9268-7-1","citationCount":"16","resultStr":"{\"title\":\"Two mosaic terminal inverted duplications arising post-zygotically: Evidence for possible formation of neo-telomeres.\",\"authors\":\"Art Daniel, Luke St Heaps, Dianne Sylvester, Sara Diaz, Gregory Peters\",\"doi\":\"10.1186/1475-9268-7-1\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p><strong>Objective: </strong>To elucidate the structure of terminal inverted duplications and to investigate potential mechanisms of formation in two cases where there was mosaicism with cells of apparently normal karyotype.</p><p><strong>Results: </strong>A karyotype [46,XY,inv dup(4)(p16.3p15.1)/46,XY] performed on blood lymphocytes from a patient referred for developmental delay (case 1) demonstrated a normal karyotype in 60% of cells with a terminal inverted duplication 4p in the remainder. In case 2, referred for multiple fetal anomalies on an ultrasound scan, 33% of amniocyte colonies were karyotypically normal, with a terminal inv dup 10p in the remainder [46,XX,inv dup(10)(p15.3p11)/46,XX]. Duplicated FISH signals for GATA3 and NEBL loci (in case 2), and for the Wolf-Hirschhorn locus (case 1) confirmed the inverted structure of both duplications. In the GTL banded normal cells from both cases, there was a cryptic deletion detected by FISH of one copy of the subtelomere 4p (case 1, probe GS-36P21), and subtelomere 10p (case 2, probe GS-306F7). At pter on both inv dup chromosomes there was no FISH signal present for the specific subtelomere probe. However, a positive pantelomeric probe signal was detected at 4 pter and 10 pter in both the cryptically-deleted chromosomes and the inv dup chromosomes in the respective cell lines of both cases.</p><p><strong>Conclusion: </strong>An inv dup structure was evident for both cases on GTL bands, and confirmed by the various FISH studies. The presence of telomere (TTAGGG repeat) sequences at pter on the inv dup chromosomes (where more proximal chromosome specific subtelomeric probes were negative) was indicated by the pantelomeric probe signals in both cases. We conclude the most likely mechanism of origin in both cases was by sub-telomeric breakage in the zygote at pter, and delayed repair/rearrangement until after one or more subsequent mitotic divisions. In these divisions, at least one breakage-fusion-bridge cycle occurred, to produce inverted duplications. It is proposed then that two differently \\\"repaired\\\" daughter cells proliferated in parallel. In one daughter cell line (with an overtly normal karyotype) there was deletion of the subtelomere and presumed repair through capping by a neo-telomere (i.e. \\\"healing\\\", as initially proposed by McClintock). This occurred in both cases presented. In the other daughter cell of each case, it is proposed that chromosome stabilization was achieved (after replication) by sister chromatid reunion to form a dicentric, which broke at a subsequent anaphase, to form an inverted duplication (with loss of the reciprocal product, and the other daughter cell line). One inv dup was repaired without an interstitial specific subtelomere (case 1) and one was repaired with a duplicated specific interstitial subtelomere (case 2). After repair TTAGGG repeats were detected by FISH at each respective new pter.</p>\",\"PeriodicalId\":84415,\"journal\":{\"name\":\"Cell & chromosome\",\"volume\":\"7 \",\"pages\":\"1\"},\"PeriodicalIF\":0.0000,\"publicationDate\":\"2008-03-10\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://sci-hub-pdf.com/10.1186/1475-9268-7-1\",\"citationCount\":\"16\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"Cell & chromosome\",\"FirstCategoryId\":\"1085\",\"ListUrlMain\":\"https://doi.org/10.1186/1475-9268-7-1\",\"RegionNum\":0,\"RegionCategory\":null,\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"\",\"JCRName\":\"\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"Cell & chromosome","FirstCategoryId":"1085","ListUrlMain":"https://doi.org/10.1186/1475-9268-7-1","RegionNum":0,"RegionCategory":null,"ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"","JCRName":"","Score":null,"Total":0}
Two mosaic terminal inverted duplications arising post-zygotically: Evidence for possible formation of neo-telomeres.
Objective: To elucidate the structure of terminal inverted duplications and to investigate potential mechanisms of formation in two cases where there was mosaicism with cells of apparently normal karyotype.
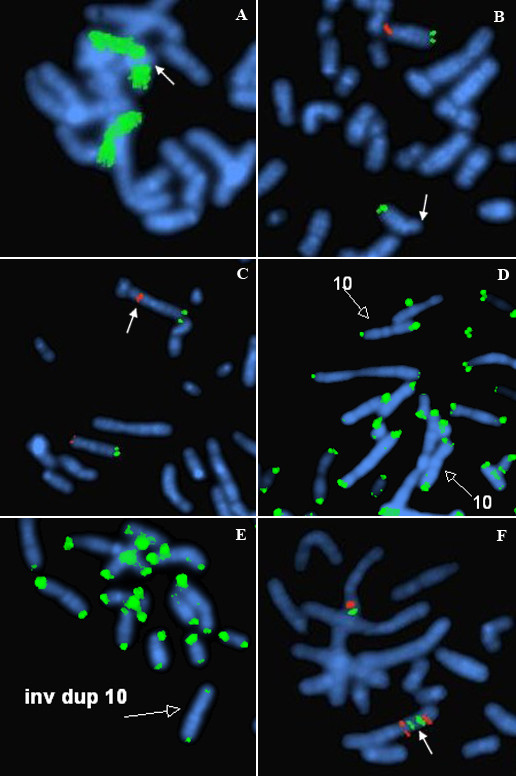
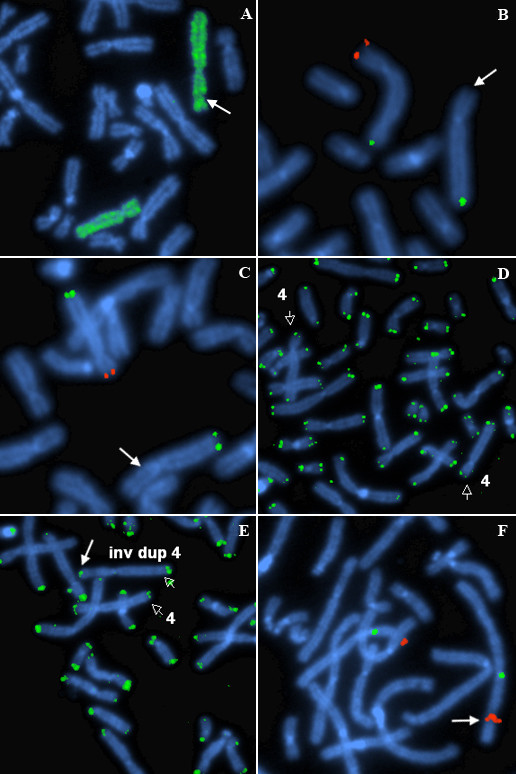
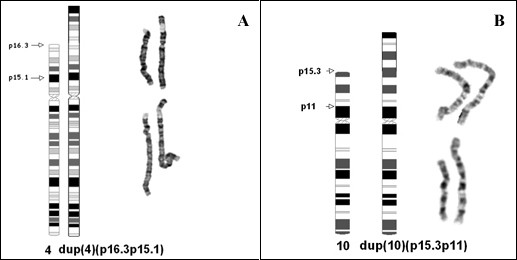
Results: A karyotype [46,XY,inv dup(4)(p16.3p15.1)/46,XY] performed on blood lymphocytes from a patient referred for developmental delay (case 1) demonstrated a normal karyotype in 60% of cells with a terminal inverted duplication 4p in the remainder. In case 2, referred for multiple fetal anomalies on an ultrasound scan, 33% of amniocyte colonies were karyotypically normal, with a terminal inv dup 10p in the remainder [46,XX,inv dup(10)(p15.3p11)/46,XX]. Duplicated FISH signals for GATA3 and NEBL loci (in case 2), and for the Wolf-Hirschhorn locus (case 1) confirmed the inverted structure of both duplications. In the GTL banded normal cells from both cases, there was a cryptic deletion detected by FISH of one copy of the subtelomere 4p (case 1, probe GS-36P21), and subtelomere 10p (case 2, probe GS-306F7). At pter on both inv dup chromosomes there was no FISH signal present for the specific subtelomere probe. However, a positive pantelomeric probe signal was detected at 4 pter and 10 pter in both the cryptically-deleted chromosomes and the inv dup chromosomes in the respective cell lines of both cases.
Conclusion: An inv dup structure was evident for both cases on GTL bands, and confirmed by the various FISH studies. The presence of telomere (TTAGGG repeat) sequences at pter on the inv dup chromosomes (where more proximal chromosome specific subtelomeric probes were negative) was indicated by the pantelomeric probe signals in both cases. We conclude the most likely mechanism of origin in both cases was by sub-telomeric breakage in the zygote at pter, and delayed repair/rearrangement until after one or more subsequent mitotic divisions. In these divisions, at least one breakage-fusion-bridge cycle occurred, to produce inverted duplications. It is proposed then that two differently "repaired" daughter cells proliferated in parallel. In one daughter cell line (with an overtly normal karyotype) there was deletion of the subtelomere and presumed repair through capping by a neo-telomere (i.e. "healing", as initially proposed by McClintock). This occurred in both cases presented. In the other daughter cell of each case, it is proposed that chromosome stabilization was achieved (after replication) by sister chromatid reunion to form a dicentric, which broke at a subsequent anaphase, to form an inverted duplication (with loss of the reciprocal product, and the other daughter cell line). One inv dup was repaired without an interstitial specific subtelomere (case 1) and one was repaired with a duplicated specific interstitial subtelomere (case 2). After repair TTAGGG repeats were detected by FISH at each respective new pter.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


