{"title":"地衣——微型温室。","authors":"Martin Grube","doi":"10.15698/mic2021.03.743","DOIUrl":null,"url":null,"abstract":"Beards hanging from trees and colorful patches encrusting rocks are silent success stories of lichens, the fascinating life styles fungi can form with algae (Fig. 1). Lichens were show-cases to introduce the concept of symbiosis (as ‘Symbiotismus’ [1]). The self-support of symbiotic life styles is recognized as gear-shift of evolution and applied to a vast number of examples where continued interactions between species lead to metabolic or phenotypic novelty. Lichen symbioses are still outstanding for the structural longevity and occurrence in environments, some which are unsuitable for most other organisms. Lichens often form major components Arctic tundra, boreal forest floors, but also on lava fields, rock surfaces along coasts or in extremely high altitudes. The perseverance of lichens in such hostile places appears to be in striking contrast to observed ecological specialization and their lack in urban and trafficated places. The symbiosis is indeed very sensitive during physiologically active state but the puzzle of extremotolerance is solved when we consider poikilohydry: because lichens hardly possess structural or functional mechanisms to maintain and/or regulate water content, desiccation rapidly causes shut down of metabolism. Yet, in contrast to many other life forms, lichens cope extremely well with recurrent changes of water availability. Lichens have an outstanding ability to revitalize from dry stages. Lichens can endure extreme desiccation to water contents (below 0.1 g H2O g–1 dry weight (DW)), which causes ‘vitrification’, the transition of their cytoplasm to a ‘glassy’ state and cease of metabolism. To find out what reactions may occur at different levels of desiccation in lichens, Candotto Carniel et al. [2] used dynamic mechanical thermal analysis as for assessment of molecular mobility, while deand re-epoxidation of the xanthophyll cycle pigments served as a proxy to assess enzyme activity. At 20°C vitrification occurred between 0.12–0.08 g H2O g−1 DW and enzymes were active in a ‘rubbery’ state (0.17 g H2O g−1 DW) but not in a glassy state (0.03 g H2O g−1 DW). Therefore, desiccated tissues may appear to be ‘dry’ in the conventional sense, but subtle differences in water content will have substantial consequences on the types of (bio)chemical reactions that can occur, with downstream effects on longevity in the desiccated state. Lichen thalli must be flexible to retain shape integrity under poikilohydric conditions, which involve shrinking and swelling of the symbiotic structures. The photosynthetic partners in the majority of lichens, algae or cyanobacteria, are typically sheltered beneath coherent peripheral layers formed by fungal cells, which are tightly glued together in a common extracellular matrix by their gelatinizing outer cell walls. Spribille et al. [3] compiled current knowledge about the composition of involved polysaccharides and emphasized the important role of acidic polysaccharides in holding lichens together. The potential effects of desiccation and rewetting (D/W) cycles for regulation of fungal polysaccharide composition still needs to be established. For the algal partner of lichens, González-Hourcade et al. [4] already discovered that exposure to D/W cycles strongly altered the size distribution of certain polysaccharides. The authors concluded that biochemical remodeling of the cell wall could increase flexibility, allowing regulated shrinkage and expansion of algal symbionts. Lack of characteristic environmental triggers, including D/W cycles, might also explain why native thallus structures lichens are hardly resynthesized in Petri dish cultures. Several attempts have recently been undertaken to achieve a better understanding of the genomic “hardwiring” for the lichen symbiosis. For example, Armaleo et al. [5] conducted a first parallel genomic analysis of the mycobiont Cladonia grayi and of its green algal photobiont Asterochloris glomerata. Gene family expansions were present in both symbionts (such as, signal transduction components, ankyrin domain proteins and transcription factors involved in chromatin remodeling and stress responses), as well as expanded fungal protein families (such as heterokaryon incompatibility proteins, polyketide synthases, and a unique set of G-protein α subunit paralogs) and expanded algal protein families (carbohydrate active enzymes and a specific subclass of cytoplasmic carbonic anhydrases). Horizontal gene transfer from prokaryotes played a likely role for acquisition of novel archaeal ATPases and Desiccation-Related Proteins by the algae. According to these results lichens evolved by accretion of many scattered regulatory and structural changes, which agrees with an independent origin of lichenized fungal lineages in the fungal","PeriodicalId":18397,"journal":{"name":"Microbial Cell","volume":"8 3","pages":"65-68"},"PeriodicalIF":3.9000,"publicationDate":"2021-03-01","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7919387/pdf/","citationCount":"2","resultStr":"{\"title\":\"Lichens - growing greenhouses <i>en miniature</i>.\",\"authors\":\"Martin Grube\",\"doi\":\"10.15698/mic2021.03.743\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"Beards hanging from trees and colorful patches encrusting rocks are silent success stories of lichens, the fascinating life styles fungi can form with algae (Fig. 1). Lichens were show-cases to introduce the concept of symbiosis (as ‘Symbiotismus’ [1]). The self-support of symbiotic life styles is recognized as gear-shift of evolution and applied to a vast number of examples where continued interactions between species lead to metabolic or phenotypic novelty. Lichen symbioses are still outstanding for the structural longevity and occurrence in environments, some which are unsuitable for most other organisms. Lichens often form major components Arctic tundra, boreal forest floors, but also on lava fields, rock surfaces along coasts or in extremely high altitudes. The perseverance of lichens in such hostile places appears to be in striking contrast to observed ecological specialization and their lack in urban and trafficated places. The symbiosis is indeed very sensitive during physiologically active state but the puzzle of extremotolerance is solved when we consider poikilohydry: because lichens hardly possess structural or functional mechanisms to maintain and/or regulate water content, desiccation rapidly causes shut down of metabolism. Yet, in contrast to many other life forms, lichens cope extremely well with recurrent changes of water availability. Lichens have an outstanding ability to revitalize from dry stages. Lichens can endure extreme desiccation to water contents (below 0.1 g H2O g–1 dry weight (DW)), which causes ‘vitrification’, the transition of their cytoplasm to a ‘glassy’ state and cease of metabolism. To find out what reactions may occur at different levels of desiccation in lichens, Candotto Carniel et al. [2] used dynamic mechanical thermal analysis as for assessment of molecular mobility, while deand re-epoxidation of the xanthophyll cycle pigments served as a proxy to assess enzyme activity. At 20°C vitrification occurred between 0.12–0.08 g H2O g−1 DW and enzymes were active in a ‘rubbery’ state (0.17 g H2O g−1 DW) but not in a glassy state (0.03 g H2O g−1 DW). Therefore, desiccated tissues may appear to be ‘dry’ in the conventional sense, but subtle differences in water content will have substantial consequences on the types of (bio)chemical reactions that can occur, with downstream effects on longevity in the desiccated state. Lichen thalli must be flexible to retain shape integrity under poikilohydric conditions, which involve shrinking and swelling of the symbiotic structures. The photosynthetic partners in the majority of lichens, algae or cyanobacteria, are typically sheltered beneath coherent peripheral layers formed by fungal cells, which are tightly glued together in a common extracellular matrix by their gelatinizing outer cell walls. Spribille et al. [3] compiled current knowledge about the composition of involved polysaccharides and emphasized the important role of acidic polysaccharides in holding lichens together. The potential effects of desiccation and rewetting (D/W) cycles for regulation of fungal polysaccharide composition still needs to be established. For the algal partner of lichens, González-Hourcade et al. [4] already discovered that exposure to D/W cycles strongly altered the size distribution of certain polysaccharides. The authors concluded that biochemical remodeling of the cell wall could increase flexibility, allowing regulated shrinkage and expansion of algal symbionts. Lack of characteristic environmental triggers, including D/W cycles, might also explain why native thallus structures lichens are hardly resynthesized in Petri dish cultures. Several attempts have recently been undertaken to achieve a better understanding of the genomic “hardwiring” for the lichen symbiosis. For example, Armaleo et al. [5] conducted a first parallel genomic analysis of the mycobiont Cladonia grayi and of its green algal photobiont Asterochloris glomerata. Gene family expansions were present in both symbionts (such as, signal transduction components, ankyrin domain proteins and transcription factors involved in chromatin remodeling and stress responses), as well as expanded fungal protein families (such as heterokaryon incompatibility proteins, polyketide synthases, and a unique set of G-protein α subunit paralogs) and expanded algal protein families (carbohydrate active enzymes and a specific subclass of cytoplasmic carbonic anhydrases). Horizontal gene transfer from prokaryotes played a likely role for acquisition of novel archaeal ATPases and Desiccation-Related Proteins by the algae. According to these results lichens evolved by accretion of many scattered regulatory and structural changes, which agrees with an independent origin of lichenized fungal lineages in the fungal\",\"PeriodicalId\":18397,\"journal\":{\"name\":\"Microbial Cell\",\"volume\":\"8 3\",\"pages\":\"65-68\"},\"PeriodicalIF\":3.9000,\"publicationDate\":\"2021-03-01\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7919387/pdf/\",\"citationCount\":\"2\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"Microbial Cell\",\"FirstCategoryId\":\"99\",\"ListUrlMain\":\"https://doi.org/10.15698/mic2021.03.743\",\"RegionNum\":3,\"RegionCategory\":\"生物学\",\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"Q2\",\"JCRName\":\"CELL BIOLOGY\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"Microbial Cell","FirstCategoryId":"99","ListUrlMain":"https://doi.org/10.15698/mic2021.03.743","RegionNum":3,"RegionCategory":"生物学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q2","JCRName":"CELL BIOLOGY","Score":null,"Total":0}
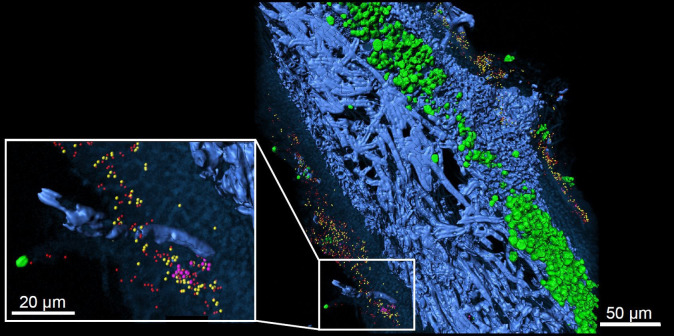
Beards hanging from trees and colorful patches encrusting rocks are silent success stories of lichens, the fascinating life styles fungi can form with algae (Fig. 1). Lichens were show-cases to introduce the concept of symbiosis (as ‘Symbiotismus’ [1]). The self-support of symbiotic life styles is recognized as gear-shift of evolution and applied to a vast number of examples where continued interactions between species lead to metabolic or phenotypic novelty. Lichen symbioses are still outstanding for the structural longevity and occurrence in environments, some which are unsuitable for most other organisms. Lichens often form major components Arctic tundra, boreal forest floors, but also on lava fields, rock surfaces along coasts or in extremely high altitudes. The perseverance of lichens in such hostile places appears to be in striking contrast to observed ecological specialization and their lack in urban and trafficated places. The symbiosis is indeed very sensitive during physiologically active state but the puzzle of extremotolerance is solved when we consider poikilohydry: because lichens hardly possess structural or functional mechanisms to maintain and/or regulate water content, desiccation rapidly causes shut down of metabolism. Yet, in contrast to many other life forms, lichens cope extremely well with recurrent changes of water availability. Lichens have an outstanding ability to revitalize from dry stages. Lichens can endure extreme desiccation to water contents (below 0.1 g H2O g–1 dry weight (DW)), which causes ‘vitrification’, the transition of their cytoplasm to a ‘glassy’ state and cease of metabolism. To find out what reactions may occur at different levels of desiccation in lichens, Candotto Carniel et al. [2] used dynamic mechanical thermal analysis as for assessment of molecular mobility, while deand re-epoxidation of the xanthophyll cycle pigments served as a proxy to assess enzyme activity. At 20°C vitrification occurred between 0.12–0.08 g H2O g−1 DW and enzymes were active in a ‘rubbery’ state (0.17 g H2O g−1 DW) but not in a glassy state (0.03 g H2O g−1 DW). Therefore, desiccated tissues may appear to be ‘dry’ in the conventional sense, but subtle differences in water content will have substantial consequences on the types of (bio)chemical reactions that can occur, with downstream effects on longevity in the desiccated state. Lichen thalli must be flexible to retain shape integrity under poikilohydric conditions, which involve shrinking and swelling of the symbiotic structures. The photosynthetic partners in the majority of lichens, algae or cyanobacteria, are typically sheltered beneath coherent peripheral layers formed by fungal cells, which are tightly glued together in a common extracellular matrix by their gelatinizing outer cell walls. Spribille et al. [3] compiled current knowledge about the composition of involved polysaccharides and emphasized the important role of acidic polysaccharides in holding lichens together. The potential effects of desiccation and rewetting (D/W) cycles for regulation of fungal polysaccharide composition still needs to be established. For the algal partner of lichens, González-Hourcade et al. [4] already discovered that exposure to D/W cycles strongly altered the size distribution of certain polysaccharides. The authors concluded that biochemical remodeling of the cell wall could increase flexibility, allowing regulated shrinkage and expansion of algal symbionts. Lack of characteristic environmental triggers, including D/W cycles, might also explain why native thallus structures lichens are hardly resynthesized in Petri dish cultures. Several attempts have recently been undertaken to achieve a better understanding of the genomic “hardwiring” for the lichen symbiosis. For example, Armaleo et al. [5] conducted a first parallel genomic analysis of the mycobiont Cladonia grayi and of its green algal photobiont Asterochloris glomerata. Gene family expansions were present in both symbionts (such as, signal transduction components, ankyrin domain proteins and transcription factors involved in chromatin remodeling and stress responses), as well as expanded fungal protein families (such as heterokaryon incompatibility proteins, polyketide synthases, and a unique set of G-protein α subunit paralogs) and expanded algal protein families (carbohydrate active enzymes and a specific subclass of cytoplasmic carbonic anhydrases). Horizontal gene transfer from prokaryotes played a likely role for acquisition of novel archaeal ATPases and Desiccation-Related Proteins by the algae. According to these results lichens evolved by accretion of many scattered regulatory and structural changes, which agrees with an independent origin of lichenized fungal lineages in the fungal


 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


