{"title":"舌头在脸颊:甜和/或鲜味的脂肪酸","authors":"Feike R. van der Leij","doi":"10.1111/apha.70088","DOIUrl":null,"url":null,"abstract":"<p>The July issue of <i>Acta Physiologica</i> contains a beautiful example of how experimental biology provides new insights into the important topic of oral reception and subsequent perception of fatty substances in mammals. In the paper “Fatty acid taste quality information via GPR40 and CD36 in the posterior tongue of mice,” Nagai and colleagues [<span>1</span>] skillfully performed surgical experiments, that, combined with additional behavioral tests shed new light on the circuitry of fatty acid signaling in the mouth. The paper contains observations that easily could have been missed if less attention would have been paid to details. The authors reach the tempting (and debatable) conclusion that long-chain fatty acids (LCFA), at least for mice, taste like sweet and/or umami tastants.</p><p>In an earlier paper by the same group, also published in Acta Physiologica [<span>2</span>], electrophysiological measurements on single chorda tympani nerve fibers coming from the anterior tongue were performed on wildtype mice and knockout mice that lack the G protein-coupled receptor GPR120, also known as free fatty acid receptor 4 (FFAR4). GPR120 is one of the proteins that have been identified about two decades ago [<span>3, 4</span>] to be involved in fatty acid tasting, together with other proteins, including the G protein-coupled receptor GPR40, also known as free fatty acid receptor 1 (FFAR1) and the LCFA transporter CD36 (“cluster of differentiation 36”) [see [<span>5</span>] for a review]. These three proteins have very diverse roles in different organs and tissues. Both GPR120 and GPR40 function in pancreatic insulin signaling, and act as the prime receptors in the gut-brain axis of fatty acid signaling that determine the long-term “wanting” of high energy nutrients like sugars and fat [<span>6</span>]. Those functions are but a few examples of many for GPR120 and GPR40. CD36, on the other hand, is the high affinity transporter needed to import the fuel into demanding tissues such as the cardiac muscle, a tissue that mainly relies on the mitochondrial oxidation of LCFA for energy generation. CD36 also has many other functions [<span>3, 5</span>].</p><p>Whether the taste of fat (by sensing of LCFA that result from oral lipase actions on triglycerides) should be considered as the sixth taste modality (next to sweet, bitter, umami, salt and sour) has long been debated, but much evidence from experimental biology pleads for it. The specific term “oleogustus” has been coined [<span>7</span>] to provide a word that is easily recognized as pertaining to the taste of oily or fatty substances without referring to other sensations of fat perception, like texture and viscosity. Indeed, humans are quite capable of tasting free fatty acids of different chain lengths. Short-chain fatty acids taste sour, medium-chain fatty acids are experienced as irritants, and LCFA taste differently than any of the other basic modalities. LCFA are described as unpalatable [<span>7</span>]. For that reason, the conclusion that mice may experience LCFA as palatable is challenging, but Nagai et al. [<span>1</span>] provide convincing evidence for LCFA activation of sweet-type and umami-type fibers in the glossopharyngeal nerve, as Yasumatsu et al. [<span>2</span>] did earlier for analogous fibers in the chorda tympani.</p><p>The electrophysiological setup applied (Figure 1A) allows for sensitive recordings of responses of dissected fibers. Sensitivity is rather important since the signals detected from LCFA stimulation of the tongue are in general much lower than those of the other tastants. Quantification of the signal patterns enables the distinction of different types of fibers. In the chorda tympani nerve fibers, those were called S-, M-, Q-, E- and N-type (sweet, umami, bitter, sour, salty, respectively) and a separate F-type (fatty acid) fiber was shown to exist. A similar typing has now been applied for fibers in the glossopharyngeal nerve, but a small change in the “alphabet” of fiber typing was needed (as the authors illustrate with a supplemental figure, the term “N-type” was replaced by “N-best” since fibers in the glossopharyngeal did not exclusively respond to NaCl as in the chorda tympani).</p><p>F-type fibers in both chorda tympani and the glossopharyngeal showed the highest response to oleic acid or linoleic acid compared to other tastants; therefore, classification of a separate F-type fiber for LCFA signaling is justified, although the prevalence was low in the chorda tympani of wildtype mice and even significantly lower in knockout mice that lack GPR120. A similar significant difference in percentages for F-type fibers has now been found for glossopharyngeal nerves, so there is some influence of GPR120 in the development of F-type circuitry, although the mechanism remains unknown for the moment. Any other involvement of GPR120 in LCFA signaling through glossopharyngeal nerve fibers was not found, which is different from what was found for chorda tympani fibers [<span>2</span>]. Earlier studies in mice showed that GPR120 is not at all needed for LCFA tasting [<span>8</span>], which matches the findings of Nagai et al. considering that in both cases taste bud cells of posterior circumvallate papillae may have initiated the LCFA signal. Judging the three studies [<span>1, 2, 8</span>] as equally valid, it illustrates the complexity of the topic as well as the influences of experimental set-up and circumstances on results.</p><p>Using specific inhibitors for GPR40 and CD36, the role of these two proteins in LCFA reception was experimentally confirmed (Figure 1B). Of course, the specificity of inhibitors should always be questioned, but the general picture is consistent with earlier results. Detailed analyses of the signaling patterns yielded a new interpretation of GPR40 and CD36 action. These kinetic differences seem more relevant for the inhibitors used than for the physiological roles of their targets (GPR40 and CD36 themselves), so this may not be the most prominent observation. However, it demonstrates the eye for details that Nagai et al. had when analyzing their results, and stimulates speculations on the mode of action of different pathways. Given the complexity of taste research, every detail counts.</p><p>The most challenging of both studies by the group of Keiko Yasumatsu, however, remains the interpretation of how LCFA are experienced by mice. Do mice, unlike men, taste LCFA really as sweet and/or umami, or does LCFA palatability depend on the strength of the solutions?</p><p>The author declares no conflicts of interest.</p>","PeriodicalId":107,"journal":{"name":"Acta Physiologica","volume":"241 9","pages":""},"PeriodicalIF":5.6000,"publicationDate":"2025-08-04","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1111/apha.70088","citationCount":"0","resultStr":"{\"title\":\"Tongue in Cheek: A Sweet and/or Umami Taste for Fatty Acids\",\"authors\":\"Feike R. van der Leij\",\"doi\":\"10.1111/apha.70088\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p>The July issue of <i>Acta Physiologica</i> contains a beautiful example of how experimental biology provides new insights into the important topic of oral reception and subsequent perception of fatty substances in mammals. In the paper “Fatty acid taste quality information via GPR40 and CD36 in the posterior tongue of mice,” Nagai and colleagues [<span>1</span>] skillfully performed surgical experiments, that, combined with additional behavioral tests shed new light on the circuitry of fatty acid signaling in the mouth. The paper contains observations that easily could have been missed if less attention would have been paid to details. The authors reach the tempting (and debatable) conclusion that long-chain fatty acids (LCFA), at least for mice, taste like sweet and/or umami tastants.</p><p>In an earlier paper by the same group, also published in Acta Physiologica [<span>2</span>], electrophysiological measurements on single chorda tympani nerve fibers coming from the anterior tongue were performed on wildtype mice and knockout mice that lack the G protein-coupled receptor GPR120, also known as free fatty acid receptor 4 (FFAR4). GPR120 is one of the proteins that have been identified about two decades ago [<span>3, 4</span>] to be involved in fatty acid tasting, together with other proteins, including the G protein-coupled receptor GPR40, also known as free fatty acid receptor 1 (FFAR1) and the LCFA transporter CD36 (“cluster of differentiation 36”) [see [<span>5</span>] for a review]. These three proteins have very diverse roles in different organs and tissues. Both GPR120 and GPR40 function in pancreatic insulin signaling, and act as the prime receptors in the gut-brain axis of fatty acid signaling that determine the long-term “wanting” of high energy nutrients like sugars and fat [<span>6</span>]. Those functions are but a few examples of many for GPR120 and GPR40. CD36, on the other hand, is the high affinity transporter needed to import the fuel into demanding tissues such as the cardiac muscle, a tissue that mainly relies on the mitochondrial oxidation of LCFA for energy generation. CD36 also has many other functions [<span>3, 5</span>].</p><p>Whether the taste of fat (by sensing of LCFA that result from oral lipase actions on triglycerides) should be considered as the sixth taste modality (next to sweet, bitter, umami, salt and sour) has long been debated, but much evidence from experimental biology pleads for it. The specific term “oleogustus” has been coined [<span>7</span>] to provide a word that is easily recognized as pertaining to the taste of oily or fatty substances without referring to other sensations of fat perception, like texture and viscosity. Indeed, humans are quite capable of tasting free fatty acids of different chain lengths. Short-chain fatty acids taste sour, medium-chain fatty acids are experienced as irritants, and LCFA taste differently than any of the other basic modalities. LCFA are described as unpalatable [<span>7</span>]. For that reason, the conclusion that mice may experience LCFA as palatable is challenging, but Nagai et al. [<span>1</span>] provide convincing evidence for LCFA activation of sweet-type and umami-type fibers in the glossopharyngeal nerve, as Yasumatsu et al. [<span>2</span>] did earlier for analogous fibers in the chorda tympani.</p><p>The electrophysiological setup applied (Figure 1A) allows for sensitive recordings of responses of dissected fibers. Sensitivity is rather important since the signals detected from LCFA stimulation of the tongue are in general much lower than those of the other tastants. Quantification of the signal patterns enables the distinction of different types of fibers. In the chorda tympani nerve fibers, those were called S-, M-, Q-, E- and N-type (sweet, umami, bitter, sour, salty, respectively) and a separate F-type (fatty acid) fiber was shown to exist. A similar typing has now been applied for fibers in the glossopharyngeal nerve, but a small change in the “alphabet” of fiber typing was needed (as the authors illustrate with a supplemental figure, the term “N-type” was replaced by “N-best” since fibers in the glossopharyngeal did not exclusively respond to NaCl as in the chorda tympani).</p><p>F-type fibers in both chorda tympani and the glossopharyngeal showed the highest response to oleic acid or linoleic acid compared to other tastants; therefore, classification of a separate F-type fiber for LCFA signaling is justified, although the prevalence was low in the chorda tympani of wildtype mice and even significantly lower in knockout mice that lack GPR120. A similar significant difference in percentages for F-type fibers has now been found for glossopharyngeal nerves, so there is some influence of GPR120 in the development of F-type circuitry, although the mechanism remains unknown for the moment. Any other involvement of GPR120 in LCFA signaling through glossopharyngeal nerve fibers was not found, which is different from what was found for chorda tympani fibers [<span>2</span>]. Earlier studies in mice showed that GPR120 is not at all needed for LCFA tasting [<span>8</span>], which matches the findings of Nagai et al. considering that in both cases taste bud cells of posterior circumvallate papillae may have initiated the LCFA signal. Judging the three studies [<span>1, 2, 8</span>] as equally valid, it illustrates the complexity of the topic as well as the influences of experimental set-up and circumstances on results.</p><p>Using specific inhibitors for GPR40 and CD36, the role of these two proteins in LCFA reception was experimentally confirmed (Figure 1B). Of course, the specificity of inhibitors should always be questioned, but the general picture is consistent with earlier results. Detailed analyses of the signaling patterns yielded a new interpretation of GPR40 and CD36 action. These kinetic differences seem more relevant for the inhibitors used than for the physiological roles of their targets (GPR40 and CD36 themselves), so this may not be the most prominent observation. However, it demonstrates the eye for details that Nagai et al. had when analyzing their results, and stimulates speculations on the mode of action of different pathways. Given the complexity of taste research, every detail counts.</p><p>The most challenging of both studies by the group of Keiko Yasumatsu, however, remains the interpretation of how LCFA are experienced by mice. Do mice, unlike men, taste LCFA really as sweet and/or umami, or does LCFA palatability depend on the strength of the solutions?</p><p>The author declares no conflicts of interest.</p>\",\"PeriodicalId\":107,\"journal\":{\"name\":\"Acta Physiologica\",\"volume\":\"241 9\",\"pages\":\"\"},\"PeriodicalIF\":5.6000,\"publicationDate\":\"2025-08-04\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://onlinelibrary.wiley.com/doi/epdf/10.1111/apha.70088\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"Acta Physiologica\",\"FirstCategoryId\":\"3\",\"ListUrlMain\":\"https://onlinelibrary.wiley.com/doi/10.1111/apha.70088\",\"RegionNum\":2,\"RegionCategory\":\"医学\",\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"Q1\",\"JCRName\":\"PHYSIOLOGY\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"Acta Physiologica","FirstCategoryId":"3","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1111/apha.70088","RegionNum":2,"RegionCategory":"医学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"PHYSIOLOGY","Score":null,"Total":0}
Tongue in Cheek: A Sweet and/or Umami Taste for Fatty Acids
The July issue of Acta Physiologica contains a beautiful example of how experimental biology provides new insights into the important topic of oral reception and subsequent perception of fatty substances in mammals. In the paper “Fatty acid taste quality information via GPR40 and CD36 in the posterior tongue of mice,” Nagai and colleagues [1] skillfully performed surgical experiments, that, combined with additional behavioral tests shed new light on the circuitry of fatty acid signaling in the mouth. The paper contains observations that easily could have been missed if less attention would have been paid to details. The authors reach the tempting (and debatable) conclusion that long-chain fatty acids (LCFA), at least for mice, taste like sweet and/or umami tastants.
In an earlier paper by the same group, also published in Acta Physiologica [2], electrophysiological measurements on single chorda tympani nerve fibers coming from the anterior tongue were performed on wildtype mice and knockout mice that lack the G protein-coupled receptor GPR120, also known as free fatty acid receptor 4 (FFAR4). GPR120 is one of the proteins that have been identified about two decades ago [3, 4] to be involved in fatty acid tasting, together with other proteins, including the G protein-coupled receptor GPR40, also known as free fatty acid receptor 1 (FFAR1) and the LCFA transporter CD36 (“cluster of differentiation 36”) [see [5] for a review]. These three proteins have very diverse roles in different organs and tissues. Both GPR120 and GPR40 function in pancreatic insulin signaling, and act as the prime receptors in the gut-brain axis of fatty acid signaling that determine the long-term “wanting” of high energy nutrients like sugars and fat [6]. Those functions are but a few examples of many for GPR120 and GPR40. CD36, on the other hand, is the high affinity transporter needed to import the fuel into demanding tissues such as the cardiac muscle, a tissue that mainly relies on the mitochondrial oxidation of LCFA for energy generation. CD36 also has many other functions [3, 5].
Whether the taste of fat (by sensing of LCFA that result from oral lipase actions on triglycerides) should be considered as the sixth taste modality (next to sweet, bitter, umami, salt and sour) has long been debated, but much evidence from experimental biology pleads for it. The specific term “oleogustus” has been coined [7] to provide a word that is easily recognized as pertaining to the taste of oily or fatty substances without referring to other sensations of fat perception, like texture and viscosity. Indeed, humans are quite capable of tasting free fatty acids of different chain lengths. Short-chain fatty acids taste sour, medium-chain fatty acids are experienced as irritants, and LCFA taste differently than any of the other basic modalities. LCFA are described as unpalatable [7]. For that reason, the conclusion that mice may experience LCFA as palatable is challenging, but Nagai et al. [1] provide convincing evidence for LCFA activation of sweet-type and umami-type fibers in the glossopharyngeal nerve, as Yasumatsu et al. [2] did earlier for analogous fibers in the chorda tympani.
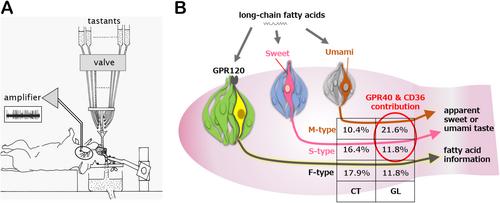
The electrophysiological setup applied (Figure 1A) allows for sensitive recordings of responses of dissected fibers. Sensitivity is rather important since the signals detected from LCFA stimulation of the tongue are in general much lower than those of the other tastants. Quantification of the signal patterns enables the distinction of different types of fibers. In the chorda tympani nerve fibers, those were called S-, M-, Q-, E- and N-type (sweet, umami, bitter, sour, salty, respectively) and a separate F-type (fatty acid) fiber was shown to exist. A similar typing has now been applied for fibers in the glossopharyngeal nerve, but a small change in the “alphabet” of fiber typing was needed (as the authors illustrate with a supplemental figure, the term “N-type” was replaced by “N-best” since fibers in the glossopharyngeal did not exclusively respond to NaCl as in the chorda tympani).
F-type fibers in both chorda tympani and the glossopharyngeal showed the highest response to oleic acid or linoleic acid compared to other tastants; therefore, classification of a separate F-type fiber for LCFA signaling is justified, although the prevalence was low in the chorda tympani of wildtype mice and even significantly lower in knockout mice that lack GPR120. A similar significant difference in percentages for F-type fibers has now been found for glossopharyngeal nerves, so there is some influence of GPR120 in the development of F-type circuitry, although the mechanism remains unknown for the moment. Any other involvement of GPR120 in LCFA signaling through glossopharyngeal nerve fibers was not found, which is different from what was found for chorda tympani fibers [2]. Earlier studies in mice showed that GPR120 is not at all needed for LCFA tasting [8], which matches the findings of Nagai et al. considering that in both cases taste bud cells of posterior circumvallate papillae may have initiated the LCFA signal. Judging the three studies [1, 2, 8] as equally valid, it illustrates the complexity of the topic as well as the influences of experimental set-up and circumstances on results.
Using specific inhibitors for GPR40 and CD36, the role of these two proteins in LCFA reception was experimentally confirmed (Figure 1B). Of course, the specificity of inhibitors should always be questioned, but the general picture is consistent with earlier results. Detailed analyses of the signaling patterns yielded a new interpretation of GPR40 and CD36 action. These kinetic differences seem more relevant for the inhibitors used than for the physiological roles of their targets (GPR40 and CD36 themselves), so this may not be the most prominent observation. However, it demonstrates the eye for details that Nagai et al. had when analyzing their results, and stimulates speculations on the mode of action of different pathways. Given the complexity of taste research, every detail counts.
The most challenging of both studies by the group of Keiko Yasumatsu, however, remains the interpretation of how LCFA are experienced by mice. Do mice, unlike men, taste LCFA really as sweet and/or umami, or does LCFA palatability depend on the strength of the solutions?
期刊介绍:
Acta Physiologica is an important forum for the publication of high quality original research in physiology and related areas by authors from all over the world. Acta Physiologica is a leading journal in human/translational physiology while promoting all aspects of the science of physiology. The journal publishes full length original articles on important new observations as well as reviews and commentaries.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


