Jacinto Román, Julio Blas, Giulia Bastianelli, Miguel A. Suárez-Couselo, Eloy Revilla, Marcello D'Amico
{"title":"道路上的殖民者:欧洲的食蜂鸟将筑巢地点的选择转移到路边的栖息地","authors":"Jacinto Román, Julio Blas, Giulia Bastianelli, Miguel A. Suárez-Couselo, Eloy Revilla, Marcello D'Amico","doi":"10.1002/ecy.70125","DOIUrl":null,"url":null,"abstract":"<p>Nest-site selection in birds results from a compromise among nest-predation risk, food availability, and microclimatic requirements (Martin, <span>1993</span>, <span>1995</span>). The European bee-eater <i>Merops apiaster</i>, an aerial-foraging insectivorous bird, breeds gregariously by digging tunnel nests at a sufficient depth underground to mitigate the impact of surface temperature (Casas-Crivillé & Valera, <span>2005</span>; White et al., <span>1978</span>). Additionally, substrate quality is crucial, requiring soils that are soft enough to be excavated yet firm enough to prevent nest collapse (Heneberg & Šimeček, <span>2004</span>; White et al., <span>1978</span>).</p><p>Doñana is located in the SW of the Iberian Peninsula (Figure 1a) and is mainly composed of two large flat environments: a seasonal freshwater marshland on clayey soils and an extensive system of eolian dune fields hosting Mediterranean woodlands and scrublands (Valverde, <span>1963</span>). This area includes Doñana National Park, where the European bee-eater has historically been a common nester along the ecotone between marshes and eolian dune fields (locally known as <i>La Vera</i>), on sandy islands within the marshes, and occasionally in other locations, such as the bottom of dry seasonal ponds (Valverde, <span>1960</span>; Figure 1b). In all these environments, the mixture of sandy and clay soils provides a suitable substrate for nest excavation. Historical records in the field notebooks of Doñana's ornithologists report bee-eater breeding colonies in the National Park and two Special Protection Areas (known as <i>El Acebuche</i> and <i>La Rocina</i>). During the 30-year period between 1970 and 1999, 17 colonies (ca. 77% of the colonies) and 1543 nests (ca. 87%) were recorded in the ecotone. Interestingly, only one colony was reported in 1993 on a roadside in <i>La Rocina</i>, with approximately 40 nests, representing 2% of the nests recorded in the historical data set (Figures 1b and 2).</p><p>Only more recently (in the year 2000), 56 nests were reported for the first time along the verges of the Palace Road, a gravel road leading to the Palace of Doñana, and bee-eaters have continued to breed there every year since then (Figures 1b–d and 3). In the following years, bee-eaters also started nesting increasingly along the verges of other roads, many outside the National Park (Figure 1e). Nesting on roadsides is not entirely novel for this species; however, most records in the scientific literature refer to anecdotal observations (Gyurácz et al., <span>2013</span>; Urbán et al., <span>2013</span>). The aim of this note is to describe this change in nest-site selection, suggest possible causes, and discuss the implications for bee-eater populations.</p><p>In 2011, 2016, and 2022, we carried out three complete censuses of bee-eater active nests across the ecotone and the eolian dune fields (Figure 1c–e). Within the National Park and the Special Protection Areas, in 2011, we recorded 327 roadside nests from a total of 533 nests counted for the whole area (ca. 61%), and in 2016, we documented 190 roadside nests from a total of 202 nests (ca. 94%), and in 2022 we identified 546 roadside nests from a total of 561 nests (ca. 97%; Figures 1c–e and 2). Additionally, in those same years, we counted a total of 357 nests outside the National Park and the Special Protection Areas, of which 343 (ca. 96%) were located along roads (note that we do not have historical data outside the park, Figure 1e).</p><p>Most roads in the region were built between the late 1960s and early 1970s, and thus, they have been available as nesting sites since the beginning of the data series collected in the Field notebooks of Doñana's ornithologists. However, European bee-eaters did not start breeding on the Palace Road until 2000, coinciding with an increase in the local livestock pressure in the ecotone, which had been the main historical breeding area for bee-eaters. In Doñana, the absence of topographic slopes implies that European bee-eaters excavate their nests directly in the ground, which are easily collapsed when trampled by livestock, including both free-ranging horses <i>Equus caballus</i> and cattle <i>Bos taurus</i>. Nest trampling by livestock is a well-documented cause of breeding failure in European bee-eaters (Arbeiter et al., <span>2014</span>; Thiollay, <span>2006</span>), and we have observed this issue in our study area as well. In Doñana, different areas of the National Park are divided by livestock fences (Botting et al., <span>2023</span>), and livestock density is artificially managed within these areas (Soriguer et al., <span>2001</span>). In the ecotone area, this increase was remarkable in the years leading up to 2000, with the annual mean number of cattle rising 3.3 times over a short time period: from an average of 293 individuals (SD = 83) in 1982–1986 to 973 (SD = 185) in 1995–1999, peaking at 1203 in 1996 (Unpublished data from Livestock Management Plan of Doñana). This high density has persisted in recent years. However, such an increase in livestock did not occur in the area around the Palace Road (mean = 94, SD = 19 in 1982–1986; mean = 95, SD = 30 in 1995–1999). This difference in livestock pressure between the ecotone and the area around the Palace Road, along with the potentially differing rates in nesting failure due to trampling, could underlie the reported change in nest-site selection by local bee-eaters. On roadsides, the mixture of loose eolian sand with more compact road-construction materials provided a very suitable substrate for nest excavation by bee-eaters in an otherwise sandy environment. In addition, large herbivores in Doñana have been reported to avoid roads consistently (D'Amico et al., <span>2016</span>; Suárez-Esteban et al., <span>2014</span>).</p><p>Additionally, another potential factor favoring this shift in nest-site selection may be the difference in predation pressure between roadsides and natural habitats. In Doñana, we have observed that the main predators of bee-eater nests are primarily the European badger <i>Meles meles</i>, but also the red fox <i>Vulpes vulpes</i> and the large gray mongoose <i>Herpestes ichneumon</i>. At least the first two species have been described as common predators of bee-eaters in other regions (Gyurácz et al., <span>2013</span>), and we know that, at least in Doñana, badgers tend to avoid roads (Revilla et al., <span>2001</span>; Suárez-Esteban et al., <span>2013</span>).</p><p>Considering that nest failure is a powerful selective pressure on bird breeding habits (Blackmer et al., <span>2004</span>; Martin, <span>1995</span>) and that previous experience, along with the success of conspecifics, serves as a key cue for nest-site selection in subsequent years (Danchin et al., <span>1998</span>; Hatchwell et al., <span>1999</span>; Ibáñez-Álamo et al., <span>2015</span>), this factor may have promoted the expansion of bee-eaters into other roads of the region. In addition, natal philopatry has been described in this species (Lessells et al., <span>1994</span>; Lessells & Krebs, <span>1989</span>) and may have contributed to the consolidation of roadside-nesting behavior.</p><p>Interestingly, European bee-eaters have not just started nesting on every roadside in our study area. In Doñana National Park, there are more than 1100 km of sandy roads with a traffic intensity of less than one vehicle/day, whereas there are only a few gravel or paved roads, all of them with medium or high traffic volumes (i.e., ca. 50–400 and >3000 vehicles/day, respectively; Román et al., <span>2010</span>). To date, we have only found bee-eater nests on both gravel and paved roads with medium traffic volume, but not on high-traffic paved roads or sandy roads. It is important to highlight that the local road network has remained largely unchanged during the period in which this shift in nesting-site selection has occurred (Román et al., <span>2010</span>).</p><p>Breeding on the roadside has its consequences. In a previous study (Blas et al., <span>2016</span>), we observed that roadside bee-eaters responded to the passing vehicles as if they were predators and therefore had higher frequencies of alarm calls and flock flushes than individuals nesting on the ecotone. The risk of being road-killed is also greater when nesting on the roadside and, in fact, every year several individuals are found dead in roadside colonies (Blas et al., <span>2016</span>). However, an increase of more than double in the nestling-feeding rates was observed in the roadside nests, which could result in a higher reproductive success (Blas et al., <span>2016</span>). This cost/benefit balance may have a threshold, so it is possible that this is the reason behind the absence of colonies on paved roads with high traffic volume.</p><p>Transportation infrastructure, and especially roads, plays a crucial role in shaping landscapes and connecting human populations (Laurance, <span>2015</span>; Laurance et al., <span>2014</span>). The impact of these infrastructures on wildlife, and especially bird populations, has become a topic of increasing concern in recent years (Johnson et al., <span>2022</span>; Kroeger et al., <span>2022</span>). The presence of roads alters the original state of the landscape by creating physical barriers, edge-effect zones, and modifying surrounding environments (Forman et al., <span>2003</span>; van Der Ree et al., <span>2015</span>), but it also provides nesting opportunities on roadside habitats (Morelli et al., <span>2014</span>). In this note, we reported a radical change in nest-site selection from natural environments toward roadsides. Long-term monitoring of this type of change is key to developing effective conservation strategies in order to maximize benefits and mitigate impacts of transportation infrastructure on avian communities.</p><p>Jacinto Román and Marcello D'Amico conceptualized the study. Jacinto Román led the methodology and investigation. Giulia Bastianelli, Miguel A. Suárez-Couselo, and Marcello D'Amico performed the censuses. Miguel A. Suárez-Couselo helped with the collection of historical data. Jacinto Román and Marcello D'Amico drafted the original manuscript. All authors revised and approved the final version of the manuscript. Julio Blas, Eloy Revilla, and Marcello D'Amico secured funding for the censuses.</p><p>The authors declare no conflicts of interest.</p>","PeriodicalId":11484,"journal":{"name":"Ecology","volume":"106 6","pages":""},"PeriodicalIF":4.3000,"publicationDate":"2025-06-12","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1002/ecy.70125","citationCount":"0","resultStr":"{\"title\":\"Colonizers on the road: European bee-eaters shift nest-site selection to roadside habitats\",\"authors\":\"Jacinto Román, Julio Blas, Giulia Bastianelli, Miguel A. Suárez-Couselo, Eloy Revilla, Marcello D'Amico\",\"doi\":\"10.1002/ecy.70125\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p>Nest-site selection in birds results from a compromise among nest-predation risk, food availability, and microclimatic requirements (Martin, <span>1993</span>, <span>1995</span>). The European bee-eater <i>Merops apiaster</i>, an aerial-foraging insectivorous bird, breeds gregariously by digging tunnel nests at a sufficient depth underground to mitigate the impact of surface temperature (Casas-Crivillé & Valera, <span>2005</span>; White et al., <span>1978</span>). Additionally, substrate quality is crucial, requiring soils that are soft enough to be excavated yet firm enough to prevent nest collapse (Heneberg & Šimeček, <span>2004</span>; White et al., <span>1978</span>).</p><p>Doñana is located in the SW of the Iberian Peninsula (Figure 1a) and is mainly composed of two large flat environments: a seasonal freshwater marshland on clayey soils and an extensive system of eolian dune fields hosting Mediterranean woodlands and scrublands (Valverde, <span>1963</span>). This area includes Doñana National Park, where the European bee-eater has historically been a common nester along the ecotone between marshes and eolian dune fields (locally known as <i>La Vera</i>), on sandy islands within the marshes, and occasionally in other locations, such as the bottom of dry seasonal ponds (Valverde, <span>1960</span>; Figure 1b). In all these environments, the mixture of sandy and clay soils provides a suitable substrate for nest excavation. Historical records in the field notebooks of Doñana's ornithologists report bee-eater breeding colonies in the National Park and two Special Protection Areas (known as <i>El Acebuche</i> and <i>La Rocina</i>). During the 30-year period between 1970 and 1999, 17 colonies (ca. 77% of the colonies) and 1543 nests (ca. 87%) were recorded in the ecotone. Interestingly, only one colony was reported in 1993 on a roadside in <i>La Rocina</i>, with approximately 40 nests, representing 2% of the nests recorded in the historical data set (Figures 1b and 2).</p><p>Only more recently (in the year 2000), 56 nests were reported for the first time along the verges of the Palace Road, a gravel road leading to the Palace of Doñana, and bee-eaters have continued to breed there every year since then (Figures 1b–d and 3). In the following years, bee-eaters also started nesting increasingly along the verges of other roads, many outside the National Park (Figure 1e). Nesting on roadsides is not entirely novel for this species; however, most records in the scientific literature refer to anecdotal observations (Gyurácz et al., <span>2013</span>; Urbán et al., <span>2013</span>). The aim of this note is to describe this change in nest-site selection, suggest possible causes, and discuss the implications for bee-eater populations.</p><p>In 2011, 2016, and 2022, we carried out three complete censuses of bee-eater active nests across the ecotone and the eolian dune fields (Figure 1c–e). Within the National Park and the Special Protection Areas, in 2011, we recorded 327 roadside nests from a total of 533 nests counted for the whole area (ca. 61%), and in 2016, we documented 190 roadside nests from a total of 202 nests (ca. 94%), and in 2022 we identified 546 roadside nests from a total of 561 nests (ca. 97%; Figures 1c–e and 2). Additionally, in those same years, we counted a total of 357 nests outside the National Park and the Special Protection Areas, of which 343 (ca. 96%) were located along roads (note that we do not have historical data outside the park, Figure 1e).</p><p>Most roads in the region were built between the late 1960s and early 1970s, and thus, they have been available as nesting sites since the beginning of the data series collected in the Field notebooks of Doñana's ornithologists. However, European bee-eaters did not start breeding on the Palace Road until 2000, coinciding with an increase in the local livestock pressure in the ecotone, which had been the main historical breeding area for bee-eaters. In Doñana, the absence of topographic slopes implies that European bee-eaters excavate their nests directly in the ground, which are easily collapsed when trampled by livestock, including both free-ranging horses <i>Equus caballus</i> and cattle <i>Bos taurus</i>. Nest trampling by livestock is a well-documented cause of breeding failure in European bee-eaters (Arbeiter et al., <span>2014</span>; Thiollay, <span>2006</span>), and we have observed this issue in our study area as well. In Doñana, different areas of the National Park are divided by livestock fences (Botting et al., <span>2023</span>), and livestock density is artificially managed within these areas (Soriguer et al., <span>2001</span>). In the ecotone area, this increase was remarkable in the years leading up to 2000, with the annual mean number of cattle rising 3.3 times over a short time period: from an average of 293 individuals (SD = 83) in 1982–1986 to 973 (SD = 185) in 1995–1999, peaking at 1203 in 1996 (Unpublished data from Livestock Management Plan of Doñana). This high density has persisted in recent years. However, such an increase in livestock did not occur in the area around the Palace Road (mean = 94, SD = 19 in 1982–1986; mean = 95, SD = 30 in 1995–1999). This difference in livestock pressure between the ecotone and the area around the Palace Road, along with the potentially differing rates in nesting failure due to trampling, could underlie the reported change in nest-site selection by local bee-eaters. On roadsides, the mixture of loose eolian sand with more compact road-construction materials provided a very suitable substrate for nest excavation by bee-eaters in an otherwise sandy environment. In addition, large herbivores in Doñana have been reported to avoid roads consistently (D'Amico et al., <span>2016</span>; Suárez-Esteban et al., <span>2014</span>).</p><p>Additionally, another potential factor favoring this shift in nest-site selection may be the difference in predation pressure between roadsides and natural habitats. In Doñana, we have observed that the main predators of bee-eater nests are primarily the European badger <i>Meles meles</i>, but also the red fox <i>Vulpes vulpes</i> and the large gray mongoose <i>Herpestes ichneumon</i>. At least the first two species have been described as common predators of bee-eaters in other regions (Gyurácz et al., <span>2013</span>), and we know that, at least in Doñana, badgers tend to avoid roads (Revilla et al., <span>2001</span>; Suárez-Esteban et al., <span>2013</span>).</p><p>Considering that nest failure is a powerful selective pressure on bird breeding habits (Blackmer et al., <span>2004</span>; Martin, <span>1995</span>) and that previous experience, along with the success of conspecifics, serves as a key cue for nest-site selection in subsequent years (Danchin et al., <span>1998</span>; Hatchwell et al., <span>1999</span>; Ibáñez-Álamo et al., <span>2015</span>), this factor may have promoted the expansion of bee-eaters into other roads of the region. In addition, natal philopatry has been described in this species (Lessells et al., <span>1994</span>; Lessells & Krebs, <span>1989</span>) and may have contributed to the consolidation of roadside-nesting behavior.</p><p>Interestingly, European bee-eaters have not just started nesting on every roadside in our study area. In Doñana National Park, there are more than 1100 km of sandy roads with a traffic intensity of less than one vehicle/day, whereas there are only a few gravel or paved roads, all of them with medium or high traffic volumes (i.e., ca. 50–400 and >3000 vehicles/day, respectively; Román et al., <span>2010</span>). To date, we have only found bee-eater nests on both gravel and paved roads with medium traffic volume, but not on high-traffic paved roads or sandy roads. It is important to highlight that the local road network has remained largely unchanged during the period in which this shift in nesting-site selection has occurred (Román et al., <span>2010</span>).</p><p>Breeding on the roadside has its consequences. In a previous study (Blas et al., <span>2016</span>), we observed that roadside bee-eaters responded to the passing vehicles as if they were predators and therefore had higher frequencies of alarm calls and flock flushes than individuals nesting on the ecotone. The risk of being road-killed is also greater when nesting on the roadside and, in fact, every year several individuals are found dead in roadside colonies (Blas et al., <span>2016</span>). However, an increase of more than double in the nestling-feeding rates was observed in the roadside nests, which could result in a higher reproductive success (Blas et al., <span>2016</span>). This cost/benefit balance may have a threshold, so it is possible that this is the reason behind the absence of colonies on paved roads with high traffic volume.</p><p>Transportation infrastructure, and especially roads, plays a crucial role in shaping landscapes and connecting human populations (Laurance, <span>2015</span>; Laurance et al., <span>2014</span>). The impact of these infrastructures on wildlife, and especially bird populations, has become a topic of increasing concern in recent years (Johnson et al., <span>2022</span>; Kroeger et al., <span>2022</span>). The presence of roads alters the original state of the landscape by creating physical barriers, edge-effect zones, and modifying surrounding environments (Forman et al., <span>2003</span>; van Der Ree et al., <span>2015</span>), but it also provides nesting opportunities on roadside habitats (Morelli et al., <span>2014</span>). In this note, we reported a radical change in nest-site selection from natural environments toward roadsides. Long-term monitoring of this type of change is key to developing effective conservation strategies in order to maximize benefits and mitigate impacts of transportation infrastructure on avian communities.</p><p>Jacinto Román and Marcello D'Amico conceptualized the study. Jacinto Román led the methodology and investigation. Giulia Bastianelli, Miguel A. Suárez-Couselo, and Marcello D'Amico performed the censuses. Miguel A. Suárez-Couselo helped with the collection of historical data. Jacinto Román and Marcello D'Amico drafted the original manuscript. All authors revised and approved the final version of the manuscript. Julio Blas, Eloy Revilla, and Marcello D'Amico secured funding for the censuses.</p><p>The authors declare no conflicts of interest.</p>\",\"PeriodicalId\":11484,\"journal\":{\"name\":\"Ecology\",\"volume\":\"106 6\",\"pages\":\"\"},\"PeriodicalIF\":4.3000,\"publicationDate\":\"2025-06-12\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://onlinelibrary.wiley.com/doi/epdf/10.1002/ecy.70125\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"Ecology\",\"FirstCategoryId\":\"93\",\"ListUrlMain\":\"https://esajournals.onlinelibrary.wiley.com/doi/10.1002/ecy.70125\",\"RegionNum\":2,\"RegionCategory\":\"环境科学与生态学\",\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"Q1\",\"JCRName\":\"ECOLOGY\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"Ecology","FirstCategoryId":"93","ListUrlMain":"https://esajournals.onlinelibrary.wiley.com/doi/10.1002/ecy.70125","RegionNum":2,"RegionCategory":"环境科学与生态学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"ECOLOGY","Score":null,"Total":0}
引用次数: 0
摘要
鸟类的筑巢地点选择是在捕食风险、食物可得性和小气候要求之间进行折衷的结果(Martin, 1993,1995)。欧洲食蜂鸟,一种空中觅食的食虫鸟,群居繁殖,在地下足够深的地方挖洞筑巢,以减轻地表温度的影响(casas - crivill& amp;瓦勒拉,2005;White et al., 1978)。此外,基材质量也至关重要,要求土壤足够软,可以挖掘,但又足够坚固,以防止巢倒塌(Heneberg &;Šimeček, 2004;White et al., 1978)。Doñana位于伊比利亚半岛的西南部(图1a),主要由两个大的平坦环境组成:一个是粘土上的季节性淡水沼泽地,一个是拥有地中海林地和灌木丛的广泛风成沙丘田系统(Valverde, 1963)。这个区域包括Doñana国家公园,欧洲食蜂鸟在历史上一直是沿着沼泽和风成沙丘地带(当地称为La Vera)之间的过渡带,在沼泽内的沙岛上,偶尔在其他地方筑巢,如干旱季节池塘的底部(Valverde, 1960;图1 b)。在所有这些环境中,砂质和粘土的混合物为挖掘巢穴提供了合适的基质。Doñana鸟类学家的野外笔记中的历史记录显示,在国家公园和两个特别保护区(称为El Acebuche和La Rocina)有食蜂鸟的繁殖群。在1970年至1999年的30年间,在过渡带记录了17个蜂群(约占蜂群的77%)和1543个巢穴(约占87%)。有趣的是,1993年在La Rocina的路边只报告了一个蚁群,大约有40个巢,占历史数据集中记录的巢的2%(图1b和2)。直到最近(2000年),第一次在宫殿路(一条通往Doñana宫殿的砾石路)的边缘发现了56个巢,从那时起,食蜂鸟每年都在那里繁殖(图1b-d和3)。在接下来的几年里,食蜂鸟也开始越来越多地在其他道路的边缘筑巢,许多都在国家公园之外(图1e)。对这个物种来说,在路边筑巢并不完全是新鲜事;然而,科学文献中的大多数记录都是轶事观察(Gyurácz et al., 2013;Urbán et al., 2013)。本文的目的是描述巢址选择的这种变化,提出可能的原因,并讨论对食蜂鸟种群的影响。在2011年、2016年和2022年,我们对整个过渡带和风成沙丘地区的食蜂鸟活跃巢穴进行了三次完整的普查(图1c-e)。在国家公园和特殊保护区,2011年,我们从全区域533个巢中记录了327个巢(约占61%),2016年,我们从202个巢中记录了190个巢(约占94%),2022年,我们从561个巢中发现了546个巢(约占97%);图1c-e和2)。此外,在同一年,我们统计了在国家公园和特别保护区外共有357个鸟巢,其中343个(约96%)位于公路沿线(注意,我们没有公园外的历史数据,图1e)。该地区的大多数道路建于20世纪60年代末至70年代初,因此,从Doñana鸟类学家的野外笔记中收集的一系列数据开始,它们就成为了筑巢地。然而,欧洲食蜂鸟直到2000年才开始在皇宫路繁殖,与此同时,过渡带当地牲畜的压力也在增加,这是历史上主要的食蜂鸟繁殖区。在Doñana,地形坡度的缺乏意味着欧洲的食蜂鸟直接在地面上挖巢,当被牲畜踩踏时很容易倒塌,包括自由放养的马Equus caballus和牛Bos taurus。牲畜践踏巢穴是欧洲食蜂鸟繁殖失败的一个有充分证据的原因(Arbeiter等人,2014;Thiollay, 2006),我们在我们的研究领域也观察到了这个问题。在Doñana中,国家公园的不同区域被牲畜围栏隔开(Botting等人,2023年),这些区域内的牲畜密度是人工管理的(soriger等人,2001年)。在过渡带地区,这种增长在2000年之前的几年中是显著的,牛的年平均数量在短时间内增加了3.3倍:从1982-1986年的平均293头(SD = 83)增加到1995-1999年的973头(SD = 185), 1996年达到1203头的峰值(来自Doñana牲畜管理计划的未发表数据)。近年来,这种高密度一直持续着。然而,这种牲畜数量的增加并未发生在Palace Road周边地区(1982-1986年,mean = 94, SD = 19;1995-1999年平均值= 95,SD = 30)。 交替带和宫殿路周围地区之间牲畜压力的差异,以及由于践踏造成的筑巢失败率的潜在差异,可能是当地食蜂者在筑巢地点选择方面所报告的变化的基础。在路边,松散的风积砂与更致密的筑路材料的混合物为食蜂鸟在其他沙质环境中挖巢提供了非常合适的基质。此外,据报道Doñana中的大型食草动物会一直避开道路(D'Amico等人,2016;Suárez-Esteban et al., 2014)。此外,另一个有利于筑巢地点选择的潜在因素可能是路边和自然栖息地之间捕食压力的差异。在Doñana中,我们观察到食蜂鸟巢穴的主要捕食者主要是欧洲獾Meles Meles,但也有红狐Vulpes Vulpes和大型灰色猫鼬Herpestes ichneuon。在其他地区,至少前两种被描述为食蜂鸟的常见捕食者(Gyurácz等人,2013),我们知道,至少在Doñana,獾倾向于避开道路(Revilla等人,2001;Suárez-Esteban et al., 2013)。考虑到筑巢失败是鸟类繁殖习惯的强大选择压力(Blackmer et al., 2004;Martin, 1995),并且之前的经验,以及同种个体的成功,是随后几年筑巢地点选择的关键线索(Danchin et al., 1998;Hatchwell et al., 1999;Ibáñez-Álamo et al., 2015),这一因素可能促进了食蜂鸟向该地区其他道路的扩张。此外,该物种还描述了出生哲学(Lessells et al., 1994;Lessells,Krebs, 1989),并可能促成了路边筑巢行为的巩固。有趣的是,欧洲食蜂鸟并没有在我们研究区域的每条路边筑巢。在Doñana国家公园,有超过1100公里的沙质道路,交通强度小于1辆/天,而只有少数砾石或铺砌的道路,都是中等或较高的交通量(分别约为50-400辆和3000辆/天);Román et al., 2010)。到目前为止,我们只在中等交通量的砾石和铺砌道路上发现了蜂巢,而在高交通量的铺砌道路和沙质道路上没有发现。需要强调的是,在巢址选择发生转变期间,当地道路网络基本保持不变(Román et al., 2010)。在路边繁殖有其后果。在之前的一项研究中(Blas et al., 2016),我们观察到路边的食蜂鸟对过往车辆的反应就像它们是捕食者一样,因此与在过渡带筑巢的个体相比,它们发出警报和蜂拥的频率更高。当在路边筑巢时,被道路杀死的风险也更大,事实上,每年都会有几个个体在路边的殖民地中被发现死亡(Blas等人,2016)。然而,在路边的巢穴中观察到的雏鸟摄食率增加了一倍以上,这可能导致更高的繁殖成功率(Blas et al., 2016)。这种成本/收益平衡可能有一个阈值,所以这可能是高交通量的铺砌道路上没有蜂群的原因。交通基础设施,尤其是道路,在塑造景观和连接人口方面起着至关重要的作用(lawrence, 2015;lawrence et al., 2014)。近年来,这些基础设施对野生动物,尤其是鸟类种群的影响已成为人们日益关注的话题(Johnson et al., 2022;Kroeger et al., 2022)。道路的存在通过创造物理障碍、边缘效应区和改变周围环境来改变景观的原始状态(Forman et al., 2003;van Der Ree et al., 2015),但它也为路边栖息地提供筑巢机会(Morelli et al., 2014)。在这篇文章中,我们报道了从自然环境到路边筑巢地点选择的根本变化。长期监测这类变化是制定有效保护策略的关键,以最大限度地提高效益,减轻交通基础设施对鸟类群落的影响。Jacinto Román和Marcello D'Amico对这项研究进行了概念化。Jacinto Román领导了研究方法和调查。Giulia Bastianelli, Miguel A. Suárez-Couselo和Marcello D'Amico进行了人口普查。Miguel A. Suárez-Couselo帮助收集历史数据。Jacinto Román和Marcello D'Amico起草了原始手稿。所有作者修改并批准了手稿的最终版本。胡里奥·布拉斯、埃洛伊·雷维拉和马塞洛·达米科为人口普查争取到了资金。作者声明无利益冲突。
Colonizers on the road: European bee-eaters shift nest-site selection to roadside habitats
Nest-site selection in birds results from a compromise among nest-predation risk, food availability, and microclimatic requirements (Martin, 1993, 1995). The European bee-eater Merops apiaster, an aerial-foraging insectivorous bird, breeds gregariously by digging tunnel nests at a sufficient depth underground to mitigate the impact of surface temperature (Casas-Crivillé & Valera, 2005; White et al., 1978). Additionally, substrate quality is crucial, requiring soils that are soft enough to be excavated yet firm enough to prevent nest collapse (Heneberg & Šimeček, 2004; White et al., 1978).
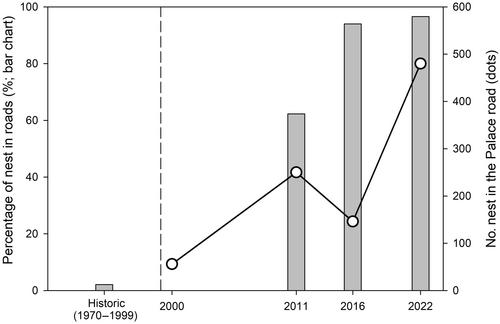
Doñana is located in the SW of the Iberian Peninsula (Figure 1a) and is mainly composed of two large flat environments: a seasonal freshwater marshland on clayey soils and an extensive system of eolian dune fields hosting Mediterranean woodlands and scrublands (Valverde, 1963). This area includes Doñana National Park, where the European bee-eater has historically been a common nester along the ecotone between marshes and eolian dune fields (locally known as La Vera), on sandy islands within the marshes, and occasionally in other locations, such as the bottom of dry seasonal ponds (Valverde, 1960; Figure 1b). In all these environments, the mixture of sandy and clay soils provides a suitable substrate for nest excavation. Historical records in the field notebooks of Doñana's ornithologists report bee-eater breeding colonies in the National Park and two Special Protection Areas (known as El Acebuche and La Rocina). During the 30-year period between 1970 and 1999, 17 colonies (ca. 77% of the colonies) and 1543 nests (ca. 87%) were recorded in the ecotone. Interestingly, only one colony was reported in 1993 on a roadside in La Rocina, with approximately 40 nests, representing 2% of the nests recorded in the historical data set (Figures 1b and 2).
Only more recently (in the year 2000), 56 nests were reported for the first time along the verges of the Palace Road, a gravel road leading to the Palace of Doñana, and bee-eaters have continued to breed there every year since then (Figures 1b–d and 3). In the following years, bee-eaters also started nesting increasingly along the verges of other roads, many outside the National Park (Figure 1e). Nesting on roadsides is not entirely novel for this species; however, most records in the scientific literature refer to anecdotal observations (Gyurácz et al., 2013; Urbán et al., 2013). The aim of this note is to describe this change in nest-site selection, suggest possible causes, and discuss the implications for bee-eater populations.
In 2011, 2016, and 2022, we carried out three complete censuses of bee-eater active nests across the ecotone and the eolian dune fields (Figure 1c–e). Within the National Park and the Special Protection Areas, in 2011, we recorded 327 roadside nests from a total of 533 nests counted for the whole area (ca. 61%), and in 2016, we documented 190 roadside nests from a total of 202 nests (ca. 94%), and in 2022 we identified 546 roadside nests from a total of 561 nests (ca. 97%; Figures 1c–e and 2). Additionally, in those same years, we counted a total of 357 nests outside the National Park and the Special Protection Areas, of which 343 (ca. 96%) were located along roads (note that we do not have historical data outside the park, Figure 1e).
Most roads in the region were built between the late 1960s and early 1970s, and thus, they have been available as nesting sites since the beginning of the data series collected in the Field notebooks of Doñana's ornithologists. However, European bee-eaters did not start breeding on the Palace Road until 2000, coinciding with an increase in the local livestock pressure in the ecotone, which had been the main historical breeding area for bee-eaters. In Doñana, the absence of topographic slopes implies that European bee-eaters excavate their nests directly in the ground, which are easily collapsed when trampled by livestock, including both free-ranging horses Equus caballus and cattle Bos taurus. Nest trampling by livestock is a well-documented cause of breeding failure in European bee-eaters (Arbeiter et al., 2014; Thiollay, 2006), and we have observed this issue in our study area as well. In Doñana, different areas of the National Park are divided by livestock fences (Botting et al., 2023), and livestock density is artificially managed within these areas (Soriguer et al., 2001). In the ecotone area, this increase was remarkable in the years leading up to 2000, with the annual mean number of cattle rising 3.3 times over a short time period: from an average of 293 individuals (SD = 83) in 1982–1986 to 973 (SD = 185) in 1995–1999, peaking at 1203 in 1996 (Unpublished data from Livestock Management Plan of Doñana). This high density has persisted in recent years. However, such an increase in livestock did not occur in the area around the Palace Road (mean = 94, SD = 19 in 1982–1986; mean = 95, SD = 30 in 1995–1999). This difference in livestock pressure between the ecotone and the area around the Palace Road, along with the potentially differing rates in nesting failure due to trampling, could underlie the reported change in nest-site selection by local bee-eaters. On roadsides, the mixture of loose eolian sand with more compact road-construction materials provided a very suitable substrate for nest excavation by bee-eaters in an otherwise sandy environment. In addition, large herbivores in Doñana have been reported to avoid roads consistently (D'Amico et al., 2016; Suárez-Esteban et al., 2014).
Additionally, another potential factor favoring this shift in nest-site selection may be the difference in predation pressure between roadsides and natural habitats. In Doñana, we have observed that the main predators of bee-eater nests are primarily the European badger Meles meles, but also the red fox Vulpes vulpes and the large gray mongoose Herpestes ichneumon. At least the first two species have been described as common predators of bee-eaters in other regions (Gyurácz et al., 2013), and we know that, at least in Doñana, badgers tend to avoid roads (Revilla et al., 2001; Suárez-Esteban et al., 2013).
Considering that nest failure is a powerful selective pressure on bird breeding habits (Blackmer et al., 2004; Martin, 1995) and that previous experience, along with the success of conspecifics, serves as a key cue for nest-site selection in subsequent years (Danchin et al., 1998; Hatchwell et al., 1999; Ibáñez-Álamo et al., 2015), this factor may have promoted the expansion of bee-eaters into other roads of the region. In addition, natal philopatry has been described in this species (Lessells et al., 1994; Lessells & Krebs, 1989) and may have contributed to the consolidation of roadside-nesting behavior.
Interestingly, European bee-eaters have not just started nesting on every roadside in our study area. In Doñana National Park, there are more than 1100 km of sandy roads with a traffic intensity of less than one vehicle/day, whereas there are only a few gravel or paved roads, all of them with medium or high traffic volumes (i.e., ca. 50–400 and >3000 vehicles/day, respectively; Román et al., 2010). To date, we have only found bee-eater nests on both gravel and paved roads with medium traffic volume, but not on high-traffic paved roads or sandy roads. It is important to highlight that the local road network has remained largely unchanged during the period in which this shift in nesting-site selection has occurred (Román et al., 2010).
Breeding on the roadside has its consequences. In a previous study (Blas et al., 2016), we observed that roadside bee-eaters responded to the passing vehicles as if they were predators and therefore had higher frequencies of alarm calls and flock flushes than individuals nesting on the ecotone. The risk of being road-killed is also greater when nesting on the roadside and, in fact, every year several individuals are found dead in roadside colonies (Blas et al., 2016). However, an increase of more than double in the nestling-feeding rates was observed in the roadside nests, which could result in a higher reproductive success (Blas et al., 2016). This cost/benefit balance may have a threshold, so it is possible that this is the reason behind the absence of colonies on paved roads with high traffic volume.
Transportation infrastructure, and especially roads, plays a crucial role in shaping landscapes and connecting human populations (Laurance, 2015; Laurance et al., 2014). The impact of these infrastructures on wildlife, and especially bird populations, has become a topic of increasing concern in recent years (Johnson et al., 2022; Kroeger et al., 2022). The presence of roads alters the original state of the landscape by creating physical barriers, edge-effect zones, and modifying surrounding environments (Forman et al., 2003; van Der Ree et al., 2015), but it also provides nesting opportunities on roadside habitats (Morelli et al., 2014). In this note, we reported a radical change in nest-site selection from natural environments toward roadsides. Long-term monitoring of this type of change is key to developing effective conservation strategies in order to maximize benefits and mitigate impacts of transportation infrastructure on avian communities.
Jacinto Román and Marcello D'Amico conceptualized the study. Jacinto Román led the methodology and investigation. Giulia Bastianelli, Miguel A. Suárez-Couselo, and Marcello D'Amico performed the censuses. Miguel A. Suárez-Couselo helped with the collection of historical data. Jacinto Román and Marcello D'Amico drafted the original manuscript. All authors revised and approved the final version of the manuscript. Julio Blas, Eloy Revilla, and Marcello D'Amico secured funding for the censuses.
期刊介绍:
Ecology publishes articles that report on the basic elements of ecological research. Emphasis is placed on concise, clear articles documenting important ecological phenomena. The journal publishes a broad array of research that includes a rapidly expanding envelope of subject matter, techniques, approaches, and concepts: paleoecology through present-day phenomena; evolutionary, population, physiological, community, and ecosystem ecology, as well as biogeochemistry; inclusive of descriptive, comparative, experimental, mathematical, statistical, and interdisciplinary approaches.



 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


