{"title":"殖民告诉我们什么?重新审视丛枝菌根真菌根定植的功能结果。","authors":"Adam Frew","doi":"10.1111/nph.70284","DOIUrl":null,"url":null,"abstract":"<p>Arbuscular mycorrhiza refers to the symbiotic association between plants and arbuscular mycorrhizal (AM) fungi, a ubiquitous and ecologically significant interaction across terrestrial ecosystems (Powell & Rillig, <span>2018</span>). In this partnership, AM fungi colonise plant roots and surrounding soil, exchanging mineral nutrients such as phosphorus for photosynthetically derived carbon from their host plants (Smith & Read, <span>2008</span>). While nutrient uptake, particularly phosphorus, is often regarded as the central functional outcome of the symbiosis, AM fungi also influence a broader suite of plant traits, including phenology, drought tolerance and defence against herbivores and pathogens (Jung <i>et al</i>., <span>2012</span>; Delavaux <i>et al</i>., <span>2017</span>; Frew <i>et al</i>., <span>2022</span>).</p><p>The physical interface of this exchange, the proportion of root length colonised by fungi, is frequently measured and used as a proxy for mycorrhizal function (McGonigle <i>et al</i>., <span>1990</span>; Smith & Smith, <span>2011</span>). This is logical; after all, it is within the roots that the symbiosis is established. Yet while it is broadly acknowledged that colonisation–benefit relationships are context dependent (Hoeksema <i>et al</i>., <span>2010</span>), this nuance often fades in practice. Perhaps because assessment of colonisation has been such a foundational and longstanding focus in mycorrhizal research, do we now interpret it with implicit assumptions of function – even when empirical support is weak or absent? While some studies report clear positive linear correlations between colonisation metrics and plant growth (Fioroni <i>et al</i>., <span>2024</span>) or phosphorus uptake (Püschel <i>et al</i>., <span>2016</span>; Ryan <i>et al</i>., <span>2016</span>), others show non-linear responses (Gange & Ayres, <span>1999</span>; Garrido <i>et al</i>., <span>2010</span>; Claassens <i>et al</i>., <span>2018</span>), or none at all (Ryan & Angus, <span>2003</span>; Ryan & Kirkegaard, <span>2012</span>; Leiser <i>et al</i>., <span>2016</span>). This raises the question of whether we continue to rely on colonisation measures out of convention, without sufficiently interrogating what they actually mean in different contexts.</p><p>Interpreting measures of colonisation is further complicated by variation among AM fungal species in their foraging strategies (although see Camenzind <i>et al</i>., <span>2024</span>) and carbon demands (Hart & Reader, <span>2002a</span>,<span>b</span>), and also by methodological limitations. The proportion of root length colonised by AM fungi (McGonigle <i>et al</i>., <span>1990</span>) is often measured at a single time point on a sub-sample of roots (understandably the assessment of an entire root system is typically not feasible), which may fail to capture the dynamic turnover of fungal structures or the spatial heterogeneity in colonisation across root types or developmental zones. Arbuscules, for example, are ephemeral structures that undergo regular turnover during the symbiosis (Toth & Miller, <span>1984</span>) meaning that their abundance at any single time point may not reflect cumulative exchange activity. Additionally, Kokkoris <i>et al</i>.'s (<span>2019</span>) valuable comparison of methods underscores that standard approaches can miss variation that may exist in the ‘intensity’ of colonisation – which incorporates the abundance of fungal structures in addition to their presence (Trouvelot, <span>1986</span>) – potentially obscuring meaningful links to function.</p><p>Despite these limitations, such measures of colonisation remain widely used as a proxy in both research and applied contexts, likely because it is relatively easy to quantify and intuitively connected to the symbiosis. However, could it also risk narrowing our expectations of what AM fungi do? Focusing exclusively on nutrient-related benefits risks underestimating other important functions of the AM symbiosis (Delavaux <i>et al</i>., <span>2017</span>). One such function is the modulation of plant defence chemistry. AM fungi are known to prime host plants for enhanced resistance to herbivores and pathogens, sometimes through the accumulation of metabolites such as phenolics (Cameron <i>et al</i>., <span>2013</span>; Pozo <i>et al</i>., <span>2013</span>; Frew, <span>2020</span>). These responses may occur with little or no corresponding change in growth or plant phosphorus (Pozo de la Hoz <i>et al</i>., <span>2021</span>; Weinberger <i>et al</i>., <span>2025</span>). Are all benefits of the symbiosis for plants mediated through exchange? Are all functions equally traceable to arbuscules?</p><p>Vannette & Hunter (<span>2011</span>) proposed a resource exchange model of defence induction, predicting that benefits such as enhanced resistance increase with the proportion of root length colonised to a point, but decline at higher colonisation levels due to carbon costs exceeding nutrient gains. Yet, it is also plausible that defence-related outcomes are initiated early in colonisation of the host by the fungi, perhaps independent of arbuscule formation or nutrient exchange (Cameron <i>et al</i>., <span>2013</span>). Defence activation from hyphal colonisation has been observed even in host species that are not arbuscular mycorrhizal (Anthony <i>et al</i>., <span>2020</span>), likely triggered by fungal surface molecules (e.g. chitin) that activate immune responses upon contact or attempted penetration (Zhang & Zhou, <span>2010</span>). Aspects of fungal colonisation can serve as cues rather than costs; if that is the case, we may be conflating structural presence and functional meaning, making it unclear what we are actually measuring.</p><p>Plant identity is another key factor shaping relationships between the proportion of root length colonised and functional outcomes. Species differ in their growth strategies, root traits and nutrient foraging behaviours, all of which can affect these colonisation-function associations (Lekberg & Koide, <span>2005</span>; Smith & Smith, <span>2011</span>; Bergmann <i>et al</i>., <span>2020</span>). C<sub>4</sub> grasses, in particular, are often considered more responsive to AM fungi, especially in terms of phosphorus uptake (Treseder, <span>2013</span>). These differences offer an opportunity to test when and where the root length colonised by the fungi serves as a meaningful proxy.</p><p>In this study, I revisit the function of colonisation metrics using a simple but structured experiment. I assessed colonisation using the McGonigle <i>et al</i>. (<span>1990</span>) intersect method and report it as the percentage of total root length colonised by fungal structures. Throughout the manuscript, I refer to the proportion of root length containing AM fungal structures (hyphae, arbuscules or vesicles) as ‘total colonisation’, and the proportion of root length colonised specifically by arbuscules as ‘arbuscular colonisation’. I measured root length colonised by AM fungi in four globally important crops (two C<sub>3</sub> and two C<sub>4</sub>) and tested their relationships to plant biomass, phosphorus and phenolics. I hypothesised that: (1) the total colonisation and arbuscular colonisation would be positively associated with plant biomass, phosphorus and phenolic responses (responses being the change in each trait in plants with AM fungi relative to controls without AM fungi), particularly in the C<sub>4</sub> crops; and (2) any relationships would be predominantly non-linear, reflecting threshold or saturation dynamics.</p><p>While measures of AM fungal colonisation remain a cornerstone metric in mycorrhizal research, their interpretation is often assumed rather than explored. This study offers empirical data across different traits and species to revisit what the root length colonised can, and cannot, tell us about symbiotic outcomes. I hope this serves as a timely prompt for deeper reflection on how we use colonisation metrics, and what we expect them to mean.</p><p>I conducted a full factorial pot experiment with four plant species that are some of the most significant crops globally: wheat (<i>Triticum aestivum</i> L. cv ‘Yitpi’), barley (<i>Hordeum vulgare</i> L. cv ‘Hindmarsh’), sorghum (<i>Sorghum bicolor</i> L. Moench cv ‘MR Taurus’) and maize (<i>Zea mays</i> L. cv ‘Amadeus’). The plants were grown in 8-l pots filled with gamma irradiated soil/sand mix (Table 1), which contained moderate to high levels of available phosphorus (46 mg kg<sup>−1</sup> Colwell P) within or above the critical phosphorus levels for these crops (see M. J. Bell <i>et al</i>., <span>2013</span>; R. Bell <i>et al</i>., <span>2013</span>), consistent with high-input crop systems. Each plant species was grown either with no AM fungi (<i>n</i> = 12 per species), or were inoculated (<i>n</i> = 24 per species) with a commercial inoculant (MicrobeSmart, Melrose Park SA) containing four species of AM fungi (<i>Entrophospora etunicatum</i> (Błaszk., B.T. Goto, Magurno, Niezgoda & Cabello) C. Walker & A. Schüßler, <i>Funneliformis coronatum</i> (Giovann.) C. Walker & A. Schüßler, <i>F. mosseae</i> (T. H. Nicolson & Gerd.) C. Walker & A. Schüßler and <i>Rhizophagus irregularis</i> (Błaszk., Wubet, Renker & Buscot) C. Walker & A. Schüßler). To encourage a broad range of fungal colonisation in roots, the AM fungal inoculant was applied at three rates – low (0.05 g kg<sup>−1</sup> soil), medium (0.5 g kg<sup>−1</sup>) and high (2 g kg<sup>−1</sup>) – with each rate given to eight plants per species, totalling 24 AM fungal-inoculated plants per species. These inoculation rates are roughly equivalent to 200, 2000 and 8000 spores per kg of soil, respectively, based on the average of 4000 spores per gram of inoculant. The inoculant was thoroughly mixed into the soil : sand substrate before potting to ensure even distribution. To account for any potential effects of non-AM fungal microbes, non-AM fungal pots were supplemented with 200 ml of microbial liquid filtrate derived from washing the AM fungal inoculum mixed with soil : sand mix filtered through sieves down to 20 μm to standardise the non-AM fungal microbial community across all pots. Plants were grown in a glasshouse with day/night temperatures of 27 and 18°C, respectively, with a 12 h photoperiod. Watering was initially 100 ml every day for the first 3 wk, followed by 400–500 ml every 3 d. Volumes were adjusted as needed to maintain consistent soil moisture across pots, monitored every 3 d using a handheld moisture meter (PMS-714; Lutron Electronic Enterprise; Taipei, Taiwan). Plants were harvested 55 d from germination, where all hosts were toward the latter vegetative growth stages, before booting or flowering. For each plant, from six locations distributed across the root system, 10–15 root fragments were collected from each location. These were cut into 5-cm segments, composited and stored in 50% ethanol before mycorrhizal fungal colonisation assessment, and the remaining aboveground and belowground plant tissues were all oven-dried at 60°C. Total biomass was recorded, and all foliar tissue was ground and homogenised before any chemical analyses.</p><p>Phosphorus (P) concentrations in plants were assessed via inductively coupled plasma spectroscopy following digestions with nitric acid (Zarcinas <i>et al</i>., <span>1987</span>). Total foliar phenolics were determined as described in Salminen & Karonen (<span>2011</span>), in technical triplicates, using a Folin–Ciocalteu assay with gallic acid monohydrate (Sigma-Aldrich, St Louis, MO, USA) as the quantification standard. For assessing root colonisation by AM fungi, and to confirm the absence of any root colonisation by AM fungi in the control plants, ethanol-stored root samples were placed into histology cassettes, then cleared with 10% potassium hydroxide (KOH) at 90°C for 10 min, stained with 5% ink-vinegar (using Quink ink; Parker Nantes, France) at 90°C for 15 min (Vierheilig <i>et al</i>., <span>1998</span>). Root fragments of 5 cm each were mounted on glass slides with glycerin under a coverslip. For each plant, 30 root fragments were assessed in total, representing 150 cm of root length. The percentage of root length colonised by any AM fungal structure (total colonisation), as well as the percentage of root length colonised by arbuscules (arbuscular colonisation), and vesicles (vesicular colonisation) were examined microscopically using the intersect method at ×200 magnification for at least 100 intersections per plant (McGonigle <i>et al</i>., <span>1990</span>). The McGonigle <i>et al</i>. (<span>1990</span>) intersect method was used as it remains the most widely applied approach (Füzy <i>et al</i>., <span>2015</span>), allowing comparability across the literature, despite the availability of alternative methods that may offer greater sensitivity (e.g. Trouvelot, <span>1986</span>; Kokkoris <i>et al</i>., <span>2019</span>).</p><p>To examine the relationship between root length colonised (total colonisation and arbuscular colonisation) and plant outcomes, plant mycorrhizal responses were calculated as ((plant response − mean plant responses with no AM fungi)/mean plant responses with no AM fungi) × 100, where the plant response was either the total biomass, P concentration, or phenolic concentration. These provided the mycorrhizal growth responses (MGR), mycorrhizal phosphorus responses (MPR) and the mycorrhizal phenolic responses (MPhenR). Due to very low vesicular colonisation across samples, vesicles were not included in the main analyses; however, these data are provided in the Supporting Information (Fig. S1c).</p><p>Generalised additive models (GAMs) were fitted for each plant species separately. These models were initially fitted for each response variable using the <span>mgcv</span> package in R (Wood, <span>2003</span>). Inoculation rates were used to generate variation in colonisation and were not included in GAMs as plant responses were analysed in relation to root length colonised, the biologically relevant predictor for the hypotheses. GAMs were chosen for their flexibility in capturing both linear and nonlinear trends without assuming a predefined functional form. Smooth terms were fitted using the default basis dimension (<i>k</i> = 10) in the <span>mgcv</span> package, the suitability of the <i>k</i> value was confirmed using diagnostic checks with the <i>gam.check</i> function. Model smoothness was determined using generalised cross-validation, and the effective degrees of freedom (edf) of the smooth term were examined to assess the shape of the relationship. Where the GAM indicated a significant relationship and edf was close to 1 (suggesting an approximately linear trend), a linear model (LM) was fitted and model selection was confirmed using Akaike Information Criterion (AIC). If a GAM indicated a significant relationship that was nonlinear (edf > 1.8), a nonlinear asymptotic model was tested using nonlinear least squares (NLS). The asymptotic model followed the self-starting asymptotic function <i>SSasymp</i> in R. Model parameters were tested for significance, and AIC values were used to confirm the best model fit. Where a GAM showed no significant relationship (<i>P</i> > 0.05 for the smooth term), no further modelling was conducted, and the response variable was considered to be independent of root length colonised. GAM fits were used to visualise relationships and the statistical outputs of the best fitting models (GAM, LM, or NLS) were reported on plots (Fig. 1). Predictions were obtained using the <i>predict</i> function over an evenly spaced sequence of colonisation values. Confidence intervals (95%) were generated from the GAM or LM and displayed as shaded regions in all plots. Where GAMs yielded non-significant relationships, no trend lines or confidence intervals were displayed in the plots. Model assumptions were checked via residual diagnostics (using the <i>gam.check</i> function for GAMs, Shapiro–Wilk tests for normality in LMs, and parameter stability checks for NLS models). Model selection was confirmed based on AIC comparisons and significance testing of fitted parameters. Statistical analyses were performed in R (v.4.3.3) using the <span>mgcv</span> (Wood, <span>2003</span>), <span>ggeffects</span> (Lüdecke, <span>2018</span>) and <span>nlme</span> packages (Pinheiro & Bates, <span>2000</span>).</p><p>Measuring the proportion of root length colonised by AM fungi offers a logical proxy for inferring the functional outcomes of the symbiosis. Yet this study adds to others that demonstrate that the relationship between colonisation and benefit is far from straightforward. It varies across traits, species and colonisation types – and, critically, not all functional outcomes appear to scale with the proportion of root length colonised in the same way.</p><p>Among all plant responses measured, the clearest and most consistent pattern emerged in relation to plant defence chemistry. In all four plant species, the total colonisation was strongly and non-linearly associated with increases in mycorrhizal phenolic responses (MPhenR), with each exhibiting an asymptotic relationship (Table S1; Fig. 1). In all species, GAMs were significant (<i>P</i> ≤ 0.01), with effective degrees of freedom (edf) ranging from 1.75 in maize to 7.56 in wheat, and high explanatory power (<i>R</i><sup>2</sup> ranging from 0.32 to 0.86). These patterns were best described by NLS models supporting my second hypothesis that colonisation–response relationships would often be non-linear, and (in part) my first, which predicted positive associations between total colonisation and phenolics.</p><p>As total colonisation increased, plant phenolic responses rose sharply at low to moderate levels, before reaching a plateau. This may suggest that phenolic defence responses are triggered by the plant signalling associated with the presence of fungi within the root (Jung <i>et al</i>., <span>2012</span>) rather than arbuscule development, and that these responses do not necessarily continue to rise in proportion to the percentage of root length colonised. That this pattern held consistently across both C<sub>3</sub> and C<sub>4</sub> crops did not support my first hypothesis that anticipated C<sub>4</sub> hosts would be more responsive.</p><p>Strikingly, no significant relationships were observed between arbuscular colonisation and MPhenR in any species (GAM <i>P</i> > 0.05; edf ≈ 1; <i>R</i><sup>2</sup> < 0.1; Table S1; Fig. 1). While some models propose that arbuscule-mediated nutrient exchange enhances defence by enabling greater resource allocation (Pozo & Azcón-Aguilar, <span>2007</span>; Vannette & Hunter, <span>2011</span>), others have demonstrated that defence responses can be activated upon fungal entry, independent of arbuscule formation or nutritional benefit (Cameron <i>et al</i>., <span>2013</span>; Anthony <i>et al</i>., <span>2020</span>; Pozo de la Hoz <i>et al</i>., <span>2021</span>). This decoupling suggests that phenolic responses are not mediated by the arbuscules themselves – despite their role as the primary interface for carbon and nutrient exchange. Arbuscular colonisation can sometimes have weak or inconsistent associations with plant traits due to the ephemeral nature of arbuscules, yet the consistent absence of any relationship across all hosts was unexpected, particularly when compared to the consistent relationship plant phenolic responses had with the total root length colonised. Rather than reflecting methodological noise due to inaccuracies of relying on measurements of the ephemeral arbuscules, this pattern may point to a biological disconnect: phenolic responses could be triggered by fungal presence more broadly, not by arbuscular function <i>per se</i>.</p><p>This distinction is important when viewed in light of the resource exchange model of plant defence from Vannette & Hunter (<span>2011</span>), which posited that defence benefits from AM fungi initially increase with colonisation but may diminish at higher fungal densities due to rising carbon costs and diminishing nutrient returns. By contrast, the patterns observed here – consistent defence responses even in the absence of growth or phosphorus benefits – suggest that the phenolic responses are not closely linked to carbon-for-nutrient exchange <i>per se</i>, as they occur even in the absence of proportional growth or phosphorus responses and are not predicted by arbuscule abundance. Rather than supporting a strict cost–benefit trade-off model, the findings here align with the understanding of a mycorrhiza-induced resistance which can be primed through early-stage fungal signalling, such as root penetration or other pre-exchange cues, independent of extensive resource exchange (Cameron <i>et al</i>., <span>2013</span>; Anthony <i>et al</i>., <span>2020</span>; Weinberger <i>et al</i>., <span>2025</span>).</p><p>The saturation of the plant phenolic responses may reflect an upper threshold of defence activation, beyond which either host regulation downregulates further metabolic investment, or fungal suppression of host defence limits further accumulation. The absence of a corresponding relationship with arbuscular colonisation does challenge the idea that nutrient exchange is required for downstream defence benefits. Instead, it suggests a broader role for colonisation as a signalling event, a cue which may then trigger systemic resistance.</p><p>Together, these findings suggest that AM fungi can modulate plant phenotype in ways that are independent of resource acquisition. Smith & Smith (<span>2011</span>) noted that P uptake via the AM fungal pathway can occur without observable growth responses, particularly in non-responsive species or under high-P conditions. Similarly, the patterns reported here imply that AM fungi may confer consistent defence-related benefits even when classical markers of mutualism (mycorrhizal growth and P responses) show little change.</p><p>In contrast to the strong phenolic patterns, and contradicting my first hypothesis, no significant associations were observed between colonisation (total or arbuscular) and MGR across any crop species (GAM <i>P</i> > 0.05; <i>R</i><sup>2</sup> ≤ 0.12). The plants in my study were grown in soil with high P availability (<i>c</i>. 46 mg kg<sup>−1</sup> Colwell P), which probably affected the likelihood of observing such effects. Nonetheless, this P level falls within the critical range reported for these crops (typically 22–48 mg kg<sup>−1</sup> depending on soil type and pH), and thus the soil P here would somewhat reflect conditions typical of high-input agricultural systems in which these crops are bred and grown. Additionally, while plant root growth was largely unrestricted, minor root binding was observed in a few sorghum replicates with particularly high root biomass – an occasional artefact of pot experiments that may influence root development or function. Although limited, such constraints could have modestly contributed to the absence of stronger growth–colonisation links. Nonetheless, average MGRs were positive across all species, with sorghum exhibiting the strongest response.</p><p>Phosphorus responses (MPR) showed clearer patterns, particularly in the C<sub>4</sub> crops. In sorghum, MPR was significantly and non-linearly associated with total colonisation (GAM edf = 1.86, <i>R</i><sup>2</sup> = 0.49, <i>P</i> < 0.001), and best fit by an NLS model. In maize, the relationship was linear (<i>R</i><sup>2</sup> = 0.35, <i>P</i> = 0.001). These findings are consistent with my first and second hypotheses and align with previous work suggesting that C<sub>4</sub> species often derive P benefits from the symbiosis in ways more tightly linked to the proportion of root length colonised (Treseder, <span>2013</span>). The asymptotic shape of the MPR response in sorghum may suggest early saturation of P benefit, while maize's linear pattern could reflect a more gradual uptake curve. These contrasting responses could challenge assumptions that P benefit may always saturate at high colonisation. Interestingly, barley showed a significant linear relationship between arbuscular colonisation and MPR (<i>R</i><sup>2</sup> = 0.41, <i>P</i> = 0.002), a result not predicted by my first or second hypothesis in that I expected the root length colonised in C<sub>4</sub> plants to have stronger relationship with MPR than in C<sub>3</sub> plants, and that this relationship would be non-linear.</p><p>Overall, the results of this study would suggest that phenolic responses may be among the most consistent outcomes of AM fungal colonisation in high-input systems, and that these may occur independently of traditional growth or nutrient-associated benefit metrics. Here I did not test whether the phenolics did confer increased resistance to pathogens or insect herbivores. This would have given a stronger basis for a functional interpretation of the MPhenR across the plant hosts.</p><p>Researchers of arbuscular mycorrhizas are well acquainted with context dependency, and the caveats here are no exception. While this study draws on data from four globally important crops, the relationships observed are likely to vary with crop variety, AM fungal identity, and environmental conditions. Rather than proposing these patterns as universal, I offer them as a prompt to revisit how we assess AM symbiosis function. This may be particularly valuable in agricultural systems, where high nutrient availability can mute growth and P mycorrhizal benefit (Ryan & Kirkegaard, <span>2012</span>).</p><p>While this study aimed to interrogate the functional meaning of colonisation metrics as they are commonly used, it is clear that methodological constraints remain a barrier to deeper understanding. Traditional staining-based techniques, including the one used here, do not discriminate between living and dead fungal structures and could conflate functional colonisation with fungal necromass. Moreover, colonisation was assessed at a single endpoint, limiting insight into the temporal dynamics of the symbiosis. Recent advances, including live-imaging techniques such as AMSlides (McGaley <i>et al</i>., <span>2025</span>) or genetically modified hosts that report arbuscule formation through anthocyanin expression (Kumar <i>et al</i>., <span>2022</span>), offer new tools to monitor colonisation over time. Similarly, molecular approaches (Bodenhausen <i>et al</i>., <span>2021</span>) targeting mitochondrial or nuclear markers could be used, and perhaps could help, to quantify metabolically active fungal tissue, while image-based analyses could allow more nuanced trait-based characterisation of intraradical structures, including potentially important distinctions in fungal morphology across host species. These emerging methods offer valuable opportunities to move beyond static, aggregate colonisation measures and to refine how we detect, interpret and ultimately understand the ecological and functional roles of AM fungal colonisation.</p><p>Finally, while the proportion of root-length colonised and similar measures remain useful and easily obtained metrics, I hope this Letter encourages careful interpretation. Indeed, the positive associations observed here between total colonisation and plant phenolics (in all four species), and between arbuscular colonisation and foliar phosphorus in three species, reinforce its utility as an informative, albeit context-dependent, indicator of symbiotic outcomes. Assessing colonisation remains fundamental to the study of the AM symbiosis; the presence of AM fungi inside the root is a prerequisite for the symbiosis itself. Yet, as we continue to rely on colonisation metrics to understand symbiotic function, we must ask not only what it measures, but what assumptions we carry with it. If we are to better understand the role of AM fungi in shaping plant traits and ecosystem processes, we must also sharpen how we interpret the most fundamental measures of the symbiosis.</p><p>The New Phytologist Foundation remains neutral with regard to jurisdictional claims in maps and in any institutional affiliations.</p>","PeriodicalId":214,"journal":{"name":"New Phytologist","volume":"247 4","pages":"1572-1578"},"PeriodicalIF":8.1000,"publicationDate":"2025-06-03","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1111/nph.70284","citationCount":"0","resultStr":"{\"title\":\"What does colonisation tell us? Revisiting the functional outcomes of root colonisation by arbuscular mycorrhizal fungi\",\"authors\":\"Adam Frew\",\"doi\":\"10.1111/nph.70284\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p>Arbuscular mycorrhiza refers to the symbiotic association between plants and arbuscular mycorrhizal (AM) fungi, a ubiquitous and ecologically significant interaction across terrestrial ecosystems (Powell & Rillig, <span>2018</span>). In this partnership, AM fungi colonise plant roots and surrounding soil, exchanging mineral nutrients such as phosphorus for photosynthetically derived carbon from their host plants (Smith & Read, <span>2008</span>). While nutrient uptake, particularly phosphorus, is often regarded as the central functional outcome of the symbiosis, AM fungi also influence a broader suite of plant traits, including phenology, drought tolerance and defence against herbivores and pathogens (Jung <i>et al</i>., <span>2012</span>; Delavaux <i>et al</i>., <span>2017</span>; Frew <i>et al</i>., <span>2022</span>).</p><p>The physical interface of this exchange, the proportion of root length colonised by fungi, is frequently measured and used as a proxy for mycorrhizal function (McGonigle <i>et al</i>., <span>1990</span>; Smith & Smith, <span>2011</span>). This is logical; after all, it is within the roots that the symbiosis is established. Yet while it is broadly acknowledged that colonisation–benefit relationships are context dependent (Hoeksema <i>et al</i>., <span>2010</span>), this nuance often fades in practice. Perhaps because assessment of colonisation has been such a foundational and longstanding focus in mycorrhizal research, do we now interpret it with implicit assumptions of function – even when empirical support is weak or absent? While some studies report clear positive linear correlations between colonisation metrics and plant growth (Fioroni <i>et al</i>., <span>2024</span>) or phosphorus uptake (Püschel <i>et al</i>., <span>2016</span>; Ryan <i>et al</i>., <span>2016</span>), others show non-linear responses (Gange & Ayres, <span>1999</span>; Garrido <i>et al</i>., <span>2010</span>; Claassens <i>et al</i>., <span>2018</span>), or none at all (Ryan & Angus, <span>2003</span>; Ryan & Kirkegaard, <span>2012</span>; Leiser <i>et al</i>., <span>2016</span>). This raises the question of whether we continue to rely on colonisation measures out of convention, without sufficiently interrogating what they actually mean in different contexts.</p><p>Interpreting measures of colonisation is further complicated by variation among AM fungal species in their foraging strategies (although see Camenzind <i>et al</i>., <span>2024</span>) and carbon demands (Hart & Reader, <span>2002a</span>,<span>b</span>), and also by methodological limitations. The proportion of root length colonised by AM fungi (McGonigle <i>et al</i>., <span>1990</span>) is often measured at a single time point on a sub-sample of roots (understandably the assessment of an entire root system is typically not feasible), which may fail to capture the dynamic turnover of fungal structures or the spatial heterogeneity in colonisation across root types or developmental zones. Arbuscules, for example, are ephemeral structures that undergo regular turnover during the symbiosis (Toth & Miller, <span>1984</span>) meaning that their abundance at any single time point may not reflect cumulative exchange activity. Additionally, Kokkoris <i>et al</i>.'s (<span>2019</span>) valuable comparison of methods underscores that standard approaches can miss variation that may exist in the ‘intensity’ of colonisation – which incorporates the abundance of fungal structures in addition to their presence (Trouvelot, <span>1986</span>) – potentially obscuring meaningful links to function.</p><p>Despite these limitations, such measures of colonisation remain widely used as a proxy in both research and applied contexts, likely because it is relatively easy to quantify and intuitively connected to the symbiosis. However, could it also risk narrowing our expectations of what AM fungi do? Focusing exclusively on nutrient-related benefits risks underestimating other important functions of the AM symbiosis (Delavaux <i>et al</i>., <span>2017</span>). One such function is the modulation of plant defence chemistry. AM fungi are known to prime host plants for enhanced resistance to herbivores and pathogens, sometimes through the accumulation of metabolites such as phenolics (Cameron <i>et al</i>., <span>2013</span>; Pozo <i>et al</i>., <span>2013</span>; Frew, <span>2020</span>). These responses may occur with little or no corresponding change in growth or plant phosphorus (Pozo de la Hoz <i>et al</i>., <span>2021</span>; Weinberger <i>et al</i>., <span>2025</span>). Are all benefits of the symbiosis for plants mediated through exchange? Are all functions equally traceable to arbuscules?</p><p>Vannette & Hunter (<span>2011</span>) proposed a resource exchange model of defence induction, predicting that benefits such as enhanced resistance increase with the proportion of root length colonised to a point, but decline at higher colonisation levels due to carbon costs exceeding nutrient gains. Yet, it is also plausible that defence-related outcomes are initiated early in colonisation of the host by the fungi, perhaps independent of arbuscule formation or nutrient exchange (Cameron <i>et al</i>., <span>2013</span>). Defence activation from hyphal colonisation has been observed even in host species that are not arbuscular mycorrhizal (Anthony <i>et al</i>., <span>2020</span>), likely triggered by fungal surface molecules (e.g. chitin) that activate immune responses upon contact or attempted penetration (Zhang & Zhou, <span>2010</span>). Aspects of fungal colonisation can serve as cues rather than costs; if that is the case, we may be conflating structural presence and functional meaning, making it unclear what we are actually measuring.</p><p>Plant identity is another key factor shaping relationships between the proportion of root length colonised and functional outcomes. Species differ in their growth strategies, root traits and nutrient foraging behaviours, all of which can affect these colonisation-function associations (Lekberg & Koide, <span>2005</span>; Smith & Smith, <span>2011</span>; Bergmann <i>et al</i>., <span>2020</span>). C<sub>4</sub> grasses, in particular, are often considered more responsive to AM fungi, especially in terms of phosphorus uptake (Treseder, <span>2013</span>). These differences offer an opportunity to test when and where the root length colonised by the fungi serves as a meaningful proxy.</p><p>In this study, I revisit the function of colonisation metrics using a simple but structured experiment. I assessed colonisation using the McGonigle <i>et al</i>. (<span>1990</span>) intersect method and report it as the percentage of total root length colonised by fungal structures. Throughout the manuscript, I refer to the proportion of root length containing AM fungal structures (hyphae, arbuscules or vesicles) as ‘total colonisation’, and the proportion of root length colonised specifically by arbuscules as ‘arbuscular colonisation’. I measured root length colonised by AM fungi in four globally important crops (two C<sub>3</sub> and two C<sub>4</sub>) and tested their relationships to plant biomass, phosphorus and phenolics. I hypothesised that: (1) the total colonisation and arbuscular colonisation would be positively associated with plant biomass, phosphorus and phenolic responses (responses being the change in each trait in plants with AM fungi relative to controls without AM fungi), particularly in the C<sub>4</sub> crops; and (2) any relationships would be predominantly non-linear, reflecting threshold or saturation dynamics.</p><p>While measures of AM fungal colonisation remain a cornerstone metric in mycorrhizal research, their interpretation is often assumed rather than explored. This study offers empirical data across different traits and species to revisit what the root length colonised can, and cannot, tell us about symbiotic outcomes. I hope this serves as a timely prompt for deeper reflection on how we use colonisation metrics, and what we expect them to mean.</p><p>I conducted a full factorial pot experiment with four plant species that are some of the most significant crops globally: wheat (<i>Triticum aestivum</i> L. cv ‘Yitpi’), barley (<i>Hordeum vulgare</i> L. cv ‘Hindmarsh’), sorghum (<i>Sorghum bicolor</i> L. Moench cv ‘MR Taurus’) and maize (<i>Zea mays</i> L. cv ‘Amadeus’). The plants were grown in 8-l pots filled with gamma irradiated soil/sand mix (Table 1), which contained moderate to high levels of available phosphorus (46 mg kg<sup>−1</sup> Colwell P) within or above the critical phosphorus levels for these crops (see M. J. Bell <i>et al</i>., <span>2013</span>; R. Bell <i>et al</i>., <span>2013</span>), consistent with high-input crop systems. Each plant species was grown either with no AM fungi (<i>n</i> = 12 per species), or were inoculated (<i>n</i> = 24 per species) with a commercial inoculant (MicrobeSmart, Melrose Park SA) containing four species of AM fungi (<i>Entrophospora etunicatum</i> (Błaszk., B.T. Goto, Magurno, Niezgoda & Cabello) C. Walker & A. Schüßler, <i>Funneliformis coronatum</i> (Giovann.) C. Walker & A. Schüßler, <i>F. mosseae</i> (T. H. Nicolson & Gerd.) C. Walker & A. Schüßler and <i>Rhizophagus irregularis</i> (Błaszk., Wubet, Renker & Buscot) C. Walker & A. Schüßler). To encourage a broad range of fungal colonisation in roots, the AM fungal inoculant was applied at three rates – low (0.05 g kg<sup>−1</sup> soil), medium (0.5 g kg<sup>−1</sup>) and high (2 g kg<sup>−1</sup>) – with each rate given to eight plants per species, totalling 24 AM fungal-inoculated plants per species. These inoculation rates are roughly equivalent to 200, 2000 and 8000 spores per kg of soil, respectively, based on the average of 4000 spores per gram of inoculant. The inoculant was thoroughly mixed into the soil : sand substrate before potting to ensure even distribution. To account for any potential effects of non-AM fungal microbes, non-AM fungal pots were supplemented with 200 ml of microbial liquid filtrate derived from washing the AM fungal inoculum mixed with soil : sand mix filtered through sieves down to 20 μm to standardise the non-AM fungal microbial community across all pots. Plants were grown in a glasshouse with day/night temperatures of 27 and 18°C, respectively, with a 12 h photoperiod. Watering was initially 100 ml every day for the first 3 wk, followed by 400–500 ml every 3 d. Volumes were adjusted as needed to maintain consistent soil moisture across pots, monitored every 3 d using a handheld moisture meter (PMS-714; Lutron Electronic Enterprise; Taipei, Taiwan). Plants were harvested 55 d from germination, where all hosts were toward the latter vegetative growth stages, before booting or flowering. For each plant, from six locations distributed across the root system, 10–15 root fragments were collected from each location. These were cut into 5-cm segments, composited and stored in 50% ethanol before mycorrhizal fungal colonisation assessment, and the remaining aboveground and belowground plant tissues were all oven-dried at 60°C. Total biomass was recorded, and all foliar tissue was ground and homogenised before any chemical analyses.</p><p>Phosphorus (P) concentrations in plants were assessed via inductively coupled plasma spectroscopy following digestions with nitric acid (Zarcinas <i>et al</i>., <span>1987</span>). Total foliar phenolics were determined as described in Salminen & Karonen (<span>2011</span>), in technical triplicates, using a Folin–Ciocalteu assay with gallic acid monohydrate (Sigma-Aldrich, St Louis, MO, USA) as the quantification standard. For assessing root colonisation by AM fungi, and to confirm the absence of any root colonisation by AM fungi in the control plants, ethanol-stored root samples were placed into histology cassettes, then cleared with 10% potassium hydroxide (KOH) at 90°C for 10 min, stained with 5% ink-vinegar (using Quink ink; Parker Nantes, France) at 90°C for 15 min (Vierheilig <i>et al</i>., <span>1998</span>). Root fragments of 5 cm each were mounted on glass slides with glycerin under a coverslip. For each plant, 30 root fragments were assessed in total, representing 150 cm of root length. The percentage of root length colonised by any AM fungal structure (total colonisation), as well as the percentage of root length colonised by arbuscules (arbuscular colonisation), and vesicles (vesicular colonisation) were examined microscopically using the intersect method at ×200 magnification for at least 100 intersections per plant (McGonigle <i>et al</i>., <span>1990</span>). The McGonigle <i>et al</i>. (<span>1990</span>) intersect method was used as it remains the most widely applied approach (Füzy <i>et al</i>., <span>2015</span>), allowing comparability across the literature, despite the availability of alternative methods that may offer greater sensitivity (e.g. Trouvelot, <span>1986</span>; Kokkoris <i>et al</i>., <span>2019</span>).</p><p>To examine the relationship between root length colonised (total colonisation and arbuscular colonisation) and plant outcomes, plant mycorrhizal responses were calculated as ((plant response − mean plant responses with no AM fungi)/mean plant responses with no AM fungi) × 100, where the plant response was either the total biomass, P concentration, or phenolic concentration. These provided the mycorrhizal growth responses (MGR), mycorrhizal phosphorus responses (MPR) and the mycorrhizal phenolic responses (MPhenR). Due to very low vesicular colonisation across samples, vesicles were not included in the main analyses; however, these data are provided in the Supporting Information (Fig. S1c).</p><p>Generalised additive models (GAMs) were fitted for each plant species separately. These models were initially fitted for each response variable using the <span>mgcv</span> package in R (Wood, <span>2003</span>). Inoculation rates were used to generate variation in colonisation and were not included in GAMs as plant responses were analysed in relation to root length colonised, the biologically relevant predictor for the hypotheses. GAMs were chosen for their flexibility in capturing both linear and nonlinear trends without assuming a predefined functional form. Smooth terms were fitted using the default basis dimension (<i>k</i> = 10) in the <span>mgcv</span> package, the suitability of the <i>k</i> value was confirmed using diagnostic checks with the <i>gam.check</i> function. Model smoothness was determined using generalised cross-validation, and the effective degrees of freedom (edf) of the smooth term were examined to assess the shape of the relationship. Where the GAM indicated a significant relationship and edf was close to 1 (suggesting an approximately linear trend), a linear model (LM) was fitted and model selection was confirmed using Akaike Information Criterion (AIC). If a GAM indicated a significant relationship that was nonlinear (edf > 1.8), a nonlinear asymptotic model was tested using nonlinear least squares (NLS). The asymptotic model followed the self-starting asymptotic function <i>SSasymp</i> in R. Model parameters were tested for significance, and AIC values were used to confirm the best model fit. Where a GAM showed no significant relationship (<i>P</i> > 0.05 for the smooth term), no further modelling was conducted, and the response variable was considered to be independent of root length colonised. GAM fits were used to visualise relationships and the statistical outputs of the best fitting models (GAM, LM, or NLS) were reported on plots (Fig. 1). Predictions were obtained using the <i>predict</i> function over an evenly spaced sequence of colonisation values. Confidence intervals (95%) were generated from the GAM or LM and displayed as shaded regions in all plots. Where GAMs yielded non-significant relationships, no trend lines or confidence intervals were displayed in the plots. Model assumptions were checked via residual diagnostics (using the <i>gam.check</i> function for GAMs, Shapiro–Wilk tests for normality in LMs, and parameter stability checks for NLS models). Model selection was confirmed based on AIC comparisons and significance testing of fitted parameters. Statistical analyses were performed in R (v.4.3.3) using the <span>mgcv</span> (Wood, <span>2003</span>), <span>ggeffects</span> (Lüdecke, <span>2018</span>) and <span>nlme</span> packages (Pinheiro & Bates, <span>2000</span>).</p><p>Measuring the proportion of root length colonised by AM fungi offers a logical proxy for inferring the functional outcomes of the symbiosis. Yet this study adds to others that demonstrate that the relationship between colonisation and benefit is far from straightforward. It varies across traits, species and colonisation types – and, critically, not all functional outcomes appear to scale with the proportion of root length colonised in the same way.</p><p>Among all plant responses measured, the clearest and most consistent pattern emerged in relation to plant defence chemistry. In all four plant species, the total colonisation was strongly and non-linearly associated with increases in mycorrhizal phenolic responses (MPhenR), with each exhibiting an asymptotic relationship (Table S1; Fig. 1). In all species, GAMs were significant (<i>P</i> ≤ 0.01), with effective degrees of freedom (edf) ranging from 1.75 in maize to 7.56 in wheat, and high explanatory power (<i>R</i><sup>2</sup> ranging from 0.32 to 0.86). These patterns were best described by NLS models supporting my second hypothesis that colonisation–response relationships would often be non-linear, and (in part) my first, which predicted positive associations between total colonisation and phenolics.</p><p>As total colonisation increased, plant phenolic responses rose sharply at low to moderate levels, before reaching a plateau. This may suggest that phenolic defence responses are triggered by the plant signalling associated with the presence of fungi within the root (Jung <i>et al</i>., <span>2012</span>) rather than arbuscule development, and that these responses do not necessarily continue to rise in proportion to the percentage of root length colonised. That this pattern held consistently across both C<sub>3</sub> and C<sub>4</sub> crops did not support my first hypothesis that anticipated C<sub>4</sub> hosts would be more responsive.</p><p>Strikingly, no significant relationships were observed between arbuscular colonisation and MPhenR in any species (GAM <i>P</i> > 0.05; edf ≈ 1; <i>R</i><sup>2</sup> < 0.1; Table S1; Fig. 1). While some models propose that arbuscule-mediated nutrient exchange enhances defence by enabling greater resource allocation (Pozo & Azcón-Aguilar, <span>2007</span>; Vannette & Hunter, <span>2011</span>), others have demonstrated that defence responses can be activated upon fungal entry, independent of arbuscule formation or nutritional benefit (Cameron <i>et al</i>., <span>2013</span>; Anthony <i>et al</i>., <span>2020</span>; Pozo de la Hoz <i>et al</i>., <span>2021</span>). This decoupling suggests that phenolic responses are not mediated by the arbuscules themselves – despite their role as the primary interface for carbon and nutrient exchange. Arbuscular colonisation can sometimes have weak or inconsistent associations with plant traits due to the ephemeral nature of arbuscules, yet the consistent absence of any relationship across all hosts was unexpected, particularly when compared to the consistent relationship plant phenolic responses had with the total root length colonised. Rather than reflecting methodological noise due to inaccuracies of relying on measurements of the ephemeral arbuscules, this pattern may point to a biological disconnect: phenolic responses could be triggered by fungal presence more broadly, not by arbuscular function <i>per se</i>.</p><p>This distinction is important when viewed in light of the resource exchange model of plant defence from Vannette & Hunter (<span>2011</span>), which posited that defence benefits from AM fungi initially increase with colonisation but may diminish at higher fungal densities due to rising carbon costs and diminishing nutrient returns. By contrast, the patterns observed here – consistent defence responses even in the absence of growth or phosphorus benefits – suggest that the phenolic responses are not closely linked to carbon-for-nutrient exchange <i>per se</i>, as they occur even in the absence of proportional growth or phosphorus responses and are not predicted by arbuscule abundance. Rather than supporting a strict cost–benefit trade-off model, the findings here align with the understanding of a mycorrhiza-induced resistance which can be primed through early-stage fungal signalling, such as root penetration or other pre-exchange cues, independent of extensive resource exchange (Cameron <i>et al</i>., <span>2013</span>; Anthony <i>et al</i>., <span>2020</span>; Weinberger <i>et al</i>., <span>2025</span>).</p><p>The saturation of the plant phenolic responses may reflect an upper threshold of defence activation, beyond which either host regulation downregulates further metabolic investment, or fungal suppression of host defence limits further accumulation. The absence of a corresponding relationship with arbuscular colonisation does challenge the idea that nutrient exchange is required for downstream defence benefits. Instead, it suggests a broader role for colonisation as a signalling event, a cue which may then trigger systemic resistance.</p><p>Together, these findings suggest that AM fungi can modulate plant phenotype in ways that are independent of resource acquisition. Smith & Smith (<span>2011</span>) noted that P uptake via the AM fungal pathway can occur without observable growth responses, particularly in non-responsive species or under high-P conditions. Similarly, the patterns reported here imply that AM fungi may confer consistent defence-related benefits even when classical markers of mutualism (mycorrhizal growth and P responses) show little change.</p><p>In contrast to the strong phenolic patterns, and contradicting my first hypothesis, no significant associations were observed between colonisation (total or arbuscular) and MGR across any crop species (GAM <i>P</i> > 0.05; <i>R</i><sup>2</sup> ≤ 0.12). The plants in my study were grown in soil with high P availability (<i>c</i>. 46 mg kg<sup>−1</sup> Colwell P), which probably affected the likelihood of observing such effects. Nonetheless, this P level falls within the critical range reported for these crops (typically 22–48 mg kg<sup>−1</sup> depending on soil type and pH), and thus the soil P here would somewhat reflect conditions typical of high-input agricultural systems in which these crops are bred and grown. Additionally, while plant root growth was largely unrestricted, minor root binding was observed in a few sorghum replicates with particularly high root biomass – an occasional artefact of pot experiments that may influence root development or function. Although limited, such constraints could have modestly contributed to the absence of stronger growth–colonisation links. Nonetheless, average MGRs were positive across all species, with sorghum exhibiting the strongest response.</p><p>Phosphorus responses (MPR) showed clearer patterns, particularly in the C<sub>4</sub> crops. In sorghum, MPR was significantly and non-linearly associated with total colonisation (GAM edf = 1.86, <i>R</i><sup>2</sup> = 0.49, <i>P</i> < 0.001), and best fit by an NLS model. In maize, the relationship was linear (<i>R</i><sup>2</sup> = 0.35, <i>P</i> = 0.001). These findings are consistent with my first and second hypotheses and align with previous work suggesting that C<sub>4</sub> species often derive P benefits from the symbiosis in ways more tightly linked to the proportion of root length colonised (Treseder, <span>2013</span>). The asymptotic shape of the MPR response in sorghum may suggest early saturation of P benefit, while maize's linear pattern could reflect a more gradual uptake curve. These contrasting responses could challenge assumptions that P benefit may always saturate at high colonisation. Interestingly, barley showed a significant linear relationship between arbuscular colonisation and MPR (<i>R</i><sup>2</sup> = 0.41, <i>P</i> = 0.002), a result not predicted by my first or second hypothesis in that I expected the root length colonised in C<sub>4</sub> plants to have stronger relationship with MPR than in C<sub>3</sub> plants, and that this relationship would be non-linear.</p><p>Overall, the results of this study would suggest that phenolic responses may be among the most consistent outcomes of AM fungal colonisation in high-input systems, and that these may occur independently of traditional growth or nutrient-associated benefit metrics. Here I did not test whether the phenolics did confer increased resistance to pathogens or insect herbivores. This would have given a stronger basis for a functional interpretation of the MPhenR across the plant hosts.</p><p>Researchers of arbuscular mycorrhizas are well acquainted with context dependency, and the caveats here are no exception. While this study draws on data from four globally important crops, the relationships observed are likely to vary with crop variety, AM fungal identity, and environmental conditions. Rather than proposing these patterns as universal, I offer them as a prompt to revisit how we assess AM symbiosis function. This may be particularly valuable in agricultural systems, where high nutrient availability can mute growth and P mycorrhizal benefit (Ryan & Kirkegaard, <span>2012</span>).</p><p>While this study aimed to interrogate the functional meaning of colonisation metrics as they are commonly used, it is clear that methodological constraints remain a barrier to deeper understanding. Traditional staining-based techniques, including the one used here, do not discriminate between living and dead fungal structures and could conflate functional colonisation with fungal necromass. Moreover, colonisation was assessed at a single endpoint, limiting insight into the temporal dynamics of the symbiosis. Recent advances, including live-imaging techniques such as AMSlides (McGaley <i>et al</i>., <span>2025</span>) or genetically modified hosts that report arbuscule formation through anthocyanin expression (Kumar <i>et al</i>., <span>2022</span>), offer new tools to monitor colonisation over time. Similarly, molecular approaches (Bodenhausen <i>et al</i>., <span>2021</span>) targeting mitochondrial or nuclear markers could be used, and perhaps could help, to quantify metabolically active fungal tissue, while image-based analyses could allow more nuanced trait-based characterisation of intraradical structures, including potentially important distinctions in fungal morphology across host species. These emerging methods offer valuable opportunities to move beyond static, aggregate colonisation measures and to refine how we detect, interpret and ultimately understand the ecological and functional roles of AM fungal colonisation.</p><p>Finally, while the proportion of root-length colonised and similar measures remain useful and easily obtained metrics, I hope this Letter encourages careful interpretation. Indeed, the positive associations observed here between total colonisation and plant phenolics (in all four species), and between arbuscular colonisation and foliar phosphorus in three species, reinforce its utility as an informative, albeit context-dependent, indicator of symbiotic outcomes. Assessing colonisation remains fundamental to the study of the AM symbiosis; the presence of AM fungi inside the root is a prerequisite for the symbiosis itself. Yet, as we continue to rely on colonisation metrics to understand symbiotic function, we must ask not only what it measures, but what assumptions we carry with it. If we are to better understand the role of AM fungi in shaping plant traits and ecosystem processes, we must also sharpen how we interpret the most fundamental measures of the symbiosis.</p><p>The New Phytologist Foundation remains neutral with regard to jurisdictional claims in maps and in any institutional affiliations.</p>\",\"PeriodicalId\":214,\"journal\":{\"name\":\"New Phytologist\",\"volume\":\"247 4\",\"pages\":\"1572-1578\"},\"PeriodicalIF\":8.1000,\"publicationDate\":\"2025-06-03\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://onlinelibrary.wiley.com/doi/epdf/10.1111/nph.70284\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"New Phytologist\",\"FirstCategoryId\":\"99\",\"ListUrlMain\":\"https://onlinelibrary.wiley.com/doi/10.1111/nph.70284\",\"RegionNum\":1,\"RegionCategory\":\"生物学\",\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"Q1\",\"JCRName\":\"PLANT SCIENCES\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"New Phytologist","FirstCategoryId":"99","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1111/nph.70284","RegionNum":1,"RegionCategory":"生物学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"PLANT SCIENCES","Score":null,"Total":0}
引用次数: 0
摘要
丛枝菌根是指植物与丛枝菌根(AM)真菌之间的共生关系,是陆地生态系统中普遍存在的具有重要生态意义的相互作用(Powell &;Rillig, 2018)。在这种合作关系中,AM真菌定植在植物根部和周围的土壤中,将磷等矿质营养物质与寄主植物的光合作用产生的碳交换(Smith &;读,2008)。虽然养分吸收,特别是磷,通常被认为是共生的核心功能结果,AM真菌也影响更广泛的植物性状,包括物候、耐旱性和对食草动物和病原体的防御(Jung等人,2012;Delavaux et al., 2017;Frew et al., 2022)。这种交换的物理界面,即真菌定殖的根长比例,经常被测量并用作菌根功能的代表(McGonigle等人,1990;史密斯,史密斯,2011)。这是合乎逻辑的;毕竟,共生关系是在根部建立起来的。然而,尽管人们普遍认为殖民-利益关系依赖于环境(Hoeksema et al., 2010),但这种细微差别在实践中往往会消失。也许是因为对菌根定植的评估一直是菌根研究的基础和长期关注的焦点,我们现在是否用隐含的功能假设来解释它——即使经验支持薄弱或缺乏?虽然一些研究报告了定植指标与植物生长(Fioroni等人,2024)或磷吸收(p<e:1>舍尔等人,2016)之间明显的正线性相关;Ryan等人,2016),其他人则表现出非线性响应(Gange &;艾尔斯,1999;Garrido et al., 2010;Claassens等人,2018),或者根本没有(Ryan &;安格斯,2003;瑞安,科克加德,2012;Leiser et al., 2016)。这就提出了一个问题,即我们是否继续依赖不合惯例的殖民措施,而没有充分询问它们在不同背景下的实际含义。AM真菌种类的觅食策略(尽管见Camenzind等人,2024)和碳需求(Hart &;Reader, 2002a,b),以及方法上的限制。AM真菌定植的根长比例(McGonigle et al., 1990)通常是在根亚样本的单个时间点上测量的(可以理解的是,对整个根系的评估通常是不可行的),这可能无法捕捉真菌结构的动态转换或不同根类型或发育区域定植的空间异质性。例如,丛枝是短暂的结构,在共生过程中经历定期的更新(Toth &;Miller, 1984),这意味着它们在任何一个时间点的丰度都不能反映累积的交换活动。此外,Kokkoris等人(2019)对方法进行的有价值的比较强调,标准方法可能会错过定殖“强度”中可能存在的变化——除了真菌结构的存在之外,还包括真菌结构的丰富度(Trouvelot, 1986)——这可能会模糊与功能的有意义的联系。尽管存在这些限制,但在研究和应用环境中,这种定殖措施仍然被广泛用作代理,可能是因为它相对容易量化并且直观地与共生联系在一起。然而,它是否也有可能缩小我们对AM真菌的预期?只关注营养相关的益处可能会低估AM共生的其他重要功能(Delavaux et al., 2017)。其中一个功能是调节植物防御化学。众所周知,AM真菌可以增强寄主植物对食草动物和病原体的抵抗力,有时通过代谢物(如酚类物质)的积累(Cameron等人,2013;Pozo et al., 2013;Frew, 2020)。这些反应可能在生长或植物磷很少或没有相应变化的情况下发生(Pozo de la Hoz等,2021;Weinberger et al., 2025)。植物共生的所有益处都是通过交换介导的吗?是否所有的功能都可以追溯到丛枝?Vannette,Hunter(2011)提出了一种防御诱导的资源交换模型,他预测,随着定殖根长度的比例达到一定程度,抗性增强等收益会增加,但在定殖水平较高时,由于碳成本超过营养收益,抗性增强等收益会下降。然而,与防御相关的结果也可能在真菌在宿主定植的早期就开始了,可能独立于丛枝形成或营养交换(Cameron et al., 2013)。即使在非丛生菌根的宿主物种中,也观察到菌丝定植的防御激活(Anthony等人,2020),可能是由真菌表面分子(例如几丁质)触发的,这些分子在接触或试图渗透时激活免疫反应(Zhang &;周,2010)。 真菌定植的各个方面可以作为线索,而不是成本;如果是这样的话,我们可能会混淆结构存在和功能意义,使我们不清楚我们实际测量的是什么。植物身份是另一个关键因素塑造的比例的根长度定殖和功能结果之间的关系。不同物种的生长策略、根系特征和营养物质觅食行为不同,所有这些都可以影响这些定殖-功能关联(Lekberg &;Koide, 2005;史密斯,史密斯,2011;Bergmann et al., 2020)。特别是C4草,通常被认为对AM真菌更敏感,特别是在磷吸收方面(Treseder, 2013)。这些差异提供了一个机会来测试真菌在何时何地定植的根长度作为一个有意义的代理。在这项研究中,我使用一个简单但结构化的实验来重新审视定植指标的功能。我使用McGonigle等人(1990)的交叉方法评估定植,并将其报告为真菌结构定植的总根长百分比。在整个手稿中,我将含有AM真菌结构(菌丝、丛枝或囊泡)的根长度比例称为“总定殖”,将丛枝具体定殖的根长度比例称为“丛枝定殖”。我测量了四种全球重要作物(两种C3和两种C4)中AM真菌定殖的根长度,并测试了它们与植物生物量、磷和酚类物质的关系。我假设:(1)总定植和丛枝定植将与植物生物量、磷和酚类反应呈正相关(反应是在有AM真菌的植物中相对于没有AM真菌的对照,每个性状的变化),特别是在C4作物中;(2)任何关系将主要是非线性的,反映阈值或饱和动态。虽然AM真菌定植的测量仍然是菌根研究的基石指标,但它们的解释往往是假设而不是探索。这项研究提供了不同性状和物种的经验数据,以重新审视定殖的根长度可以和不能告诉我们共生结果。我希望这能及时促使我们更深入地思考我们如何使用殖民指标,以及我们期望它们意味着什么。我对四种全球最重要的作物进行了全因子盆栽试验:小麦(Triticum aestivum L. cv ' Yitpi ‘)、大麦(Hordeum vulgare L. cv ’ Hindmarsh ‘)、高粱(sorghum bicolor L. Moench cv ’ MR Taurus ‘)和玉米(Zea mays L. cv ’ Amadeus ')。这些植物生长在8-l的花盆中,装满了伽马辐照的土壤/沙子混合物(表1),其中含有中高水平的有效磷(46 mg kg - 1 Colwell P),在这些作物的临界磷水平之内或以上(见M. J. Bell等人,2013;R. Bell et al., 2013),与高投入作物系统相一致。每个植物种要么没有AM真菌(n = 12 /种),要么用含有4种AM真菌(Entrophospora etunicatum (Błaszk))的商业接种剂(MicrobeSmart, Melrose Park SA)接种(n = 24 /种)。, B.T. Goto, Magurno, Niezgoda &;卡贝洛C.沃克&;A. sch<e:1> ßler,冠状漏斗状体(乔凡)C. Walker &;A. sch<e:1> ßler, F. mosseae (t.h. Nicolson &;Gerd)。C. Walker &;A. sch<s:1> ßler和不规则根噬菌(Błaszk;, Wubet, Renker &;布斯科特C.沃克&;答:Schußl)。为了促进真菌在根系中的广泛定植,AM真菌接种剂以三种速率施用-低(0.05 g kg - 1土壤),中(0.5 g kg - 1)和高(2 g kg - 1) -每种速率给予每个物种8株植物,每个物种总共接种了24株AM真菌。根据每克接种剂平均4000个孢子计算,这些接种率大致相当于每公斤土壤分别接种200、2000和8000个孢子。在灌封前将接种剂充分混入土砂基质中,确保其均匀分布。为了考虑非AM真菌微生物的任何潜在影响,在非AM真菌锅中添加200毫升微生物液滤液,这些滤液来自于将AM真菌接种物与土壤混合后的微生物液:沙子混合物通过筛子过滤至20 μm,以标准化所有锅中的非AM真菌微生物群落。植物生长在昼/夜温度分别为27°C和18°C的温室中,光周期为12 h。最初,前3周每天浇水100毫升,随后每3天浇水400-500毫升。根据需要调整水量,以保持盆间土壤湿度一致,每3天使用手持式水分计(PMS-714;路创电子企业;台北,台湾)。植株在萌发后55天收获,此时所有寄主都处于后期营养生长阶段,在抽芽或开花之前。 同样,针对线粒体或核标记的分子方法(Bodenhausen et al., 2021)可以使用,并且可能有助于量化代谢活跃的真菌组织,而基于图像的分析可以允许更细致的基于特征的根内结构特征,包括宿主物种间真菌形态的潜在重要差异。这些新兴的方法提供了宝贵的机会,超越了静态的、聚集的定植措施,并改进了我们如何检测、解释并最终理解AM真菌定植的生态和功能作用。最后,虽然根长度定植的比例和类似的度量仍然是有用的和容易获得的度量,我希望这封信鼓励仔细解释。事实上,在这里观察到的总定殖与植物酚类物质之间的正相关关系(在所有四个物种中),以及在三个物种中丛枝定殖与叶面磷之间的正相关关系,加强了它作为共生结果的信息指标的实用性,尽管依赖于环境。评估定植仍然是研究AM共生关系的基础;AM真菌在根内的存在是共生本身的先决条件。然而,当我们继续依靠殖民指标来理解共生功能时,我们不仅要问它衡量的是什么,还要问我们带有什么假设。如果我们要更好地理解AM真菌在塑造植物性状和生态系统过程中的作用,我们还必须提高我们如何解释共生关系的最基本措施。新植物学家基金会对地图和任何机构的管辖权要求保持中立。
What does colonisation tell us? Revisiting the functional outcomes of root colonisation by arbuscular mycorrhizal fungi
Arbuscular mycorrhiza refers to the symbiotic association between plants and arbuscular mycorrhizal (AM) fungi, a ubiquitous and ecologically significant interaction across terrestrial ecosystems (Powell & Rillig, 2018). In this partnership, AM fungi colonise plant roots and surrounding soil, exchanging mineral nutrients such as phosphorus for photosynthetically derived carbon from their host plants (Smith & Read, 2008). While nutrient uptake, particularly phosphorus, is often regarded as the central functional outcome of the symbiosis, AM fungi also influence a broader suite of plant traits, including phenology, drought tolerance and defence against herbivores and pathogens (Jung et al., 2012; Delavaux et al., 2017; Frew et al., 2022).
The physical interface of this exchange, the proportion of root length colonised by fungi, is frequently measured and used as a proxy for mycorrhizal function (McGonigle et al., 1990; Smith & Smith, 2011). This is logical; after all, it is within the roots that the symbiosis is established. Yet while it is broadly acknowledged that colonisation–benefit relationships are context dependent (Hoeksema et al., 2010), this nuance often fades in practice. Perhaps because assessment of colonisation has been such a foundational and longstanding focus in mycorrhizal research, do we now interpret it with implicit assumptions of function – even when empirical support is weak or absent? While some studies report clear positive linear correlations between colonisation metrics and plant growth (Fioroni et al., 2024) or phosphorus uptake (Püschel et al., 2016; Ryan et al., 2016), others show non-linear responses (Gange & Ayres, 1999; Garrido et al., 2010; Claassens et al., 2018), or none at all (Ryan & Angus, 2003; Ryan & Kirkegaard, 2012; Leiser et al., 2016). This raises the question of whether we continue to rely on colonisation measures out of convention, without sufficiently interrogating what they actually mean in different contexts.
Interpreting measures of colonisation is further complicated by variation among AM fungal species in their foraging strategies (although see Camenzind et al., 2024) and carbon demands (Hart & Reader, 2002a,b), and also by methodological limitations. The proportion of root length colonised by AM fungi (McGonigle et al., 1990) is often measured at a single time point on a sub-sample of roots (understandably the assessment of an entire root system is typically not feasible), which may fail to capture the dynamic turnover of fungal structures or the spatial heterogeneity in colonisation across root types or developmental zones. Arbuscules, for example, are ephemeral structures that undergo regular turnover during the symbiosis (Toth & Miller, 1984) meaning that their abundance at any single time point may not reflect cumulative exchange activity. Additionally, Kokkoris et al.'s (2019) valuable comparison of methods underscores that standard approaches can miss variation that may exist in the ‘intensity’ of colonisation – which incorporates the abundance of fungal structures in addition to their presence (Trouvelot, 1986) – potentially obscuring meaningful links to function.
Despite these limitations, such measures of colonisation remain widely used as a proxy in both research and applied contexts, likely because it is relatively easy to quantify and intuitively connected to the symbiosis. However, could it also risk narrowing our expectations of what AM fungi do? Focusing exclusively on nutrient-related benefits risks underestimating other important functions of the AM symbiosis (Delavaux et al., 2017). One such function is the modulation of plant defence chemistry. AM fungi are known to prime host plants for enhanced resistance to herbivores and pathogens, sometimes through the accumulation of metabolites such as phenolics (Cameron et al., 2013; Pozo et al., 2013; Frew, 2020). These responses may occur with little or no corresponding change in growth or plant phosphorus (Pozo de la Hoz et al., 2021; Weinberger et al., 2025). Are all benefits of the symbiosis for plants mediated through exchange? Are all functions equally traceable to arbuscules?
Vannette & Hunter (2011) proposed a resource exchange model of defence induction, predicting that benefits such as enhanced resistance increase with the proportion of root length colonised to a point, but decline at higher colonisation levels due to carbon costs exceeding nutrient gains. Yet, it is also plausible that defence-related outcomes are initiated early in colonisation of the host by the fungi, perhaps independent of arbuscule formation or nutrient exchange (Cameron et al., 2013). Defence activation from hyphal colonisation has been observed even in host species that are not arbuscular mycorrhizal (Anthony et al., 2020), likely triggered by fungal surface molecules (e.g. chitin) that activate immune responses upon contact or attempted penetration (Zhang & Zhou, 2010). Aspects of fungal colonisation can serve as cues rather than costs; if that is the case, we may be conflating structural presence and functional meaning, making it unclear what we are actually measuring.
Plant identity is another key factor shaping relationships between the proportion of root length colonised and functional outcomes. Species differ in their growth strategies, root traits and nutrient foraging behaviours, all of which can affect these colonisation-function associations (Lekberg & Koide, 2005; Smith & Smith, 2011; Bergmann et al., 2020). C4 grasses, in particular, are often considered more responsive to AM fungi, especially in terms of phosphorus uptake (Treseder, 2013). These differences offer an opportunity to test when and where the root length colonised by the fungi serves as a meaningful proxy.
In this study, I revisit the function of colonisation metrics using a simple but structured experiment. I assessed colonisation using the McGonigle et al. (1990) intersect method and report it as the percentage of total root length colonised by fungal structures. Throughout the manuscript, I refer to the proportion of root length containing AM fungal structures (hyphae, arbuscules or vesicles) as ‘total colonisation’, and the proportion of root length colonised specifically by arbuscules as ‘arbuscular colonisation’. I measured root length colonised by AM fungi in four globally important crops (two C3 and two C4) and tested their relationships to plant biomass, phosphorus and phenolics. I hypothesised that: (1) the total colonisation and arbuscular colonisation would be positively associated with plant biomass, phosphorus and phenolic responses (responses being the change in each trait in plants with AM fungi relative to controls without AM fungi), particularly in the C4 crops; and (2) any relationships would be predominantly non-linear, reflecting threshold or saturation dynamics.
While measures of AM fungal colonisation remain a cornerstone metric in mycorrhizal research, their interpretation is often assumed rather than explored. This study offers empirical data across different traits and species to revisit what the root length colonised can, and cannot, tell us about symbiotic outcomes. I hope this serves as a timely prompt for deeper reflection on how we use colonisation metrics, and what we expect them to mean.
I conducted a full factorial pot experiment with four plant species that are some of the most significant crops globally: wheat (Triticum aestivum L. cv ‘Yitpi’), barley (Hordeum vulgare L. cv ‘Hindmarsh’), sorghum (Sorghum bicolor L. Moench cv ‘MR Taurus’) and maize (Zea mays L. cv ‘Amadeus’). The plants were grown in 8-l pots filled with gamma irradiated soil/sand mix (Table 1), which contained moderate to high levels of available phosphorus (46 mg kg−1 Colwell P) within or above the critical phosphorus levels for these crops (see M. J. Bell et al., 2013; R. Bell et al., 2013), consistent with high-input crop systems. Each plant species was grown either with no AM fungi (n = 12 per species), or were inoculated (n = 24 per species) with a commercial inoculant (MicrobeSmart, Melrose Park SA) containing four species of AM fungi (Entrophospora etunicatum (Błaszk., B.T. Goto, Magurno, Niezgoda & Cabello) C. Walker & A. Schüßler, Funneliformis coronatum (Giovann.) C. Walker & A. Schüßler, F. mosseae (T. H. Nicolson & Gerd.) C. Walker & A. Schüßler and Rhizophagus irregularis (Błaszk., Wubet, Renker & Buscot) C. Walker & A. Schüßler). To encourage a broad range of fungal colonisation in roots, the AM fungal inoculant was applied at three rates – low (0.05 g kg−1 soil), medium (0.5 g kg−1) and high (2 g kg−1) – with each rate given to eight plants per species, totalling 24 AM fungal-inoculated plants per species. These inoculation rates are roughly equivalent to 200, 2000 and 8000 spores per kg of soil, respectively, based on the average of 4000 spores per gram of inoculant. The inoculant was thoroughly mixed into the soil : sand substrate before potting to ensure even distribution. To account for any potential effects of non-AM fungal microbes, non-AM fungal pots were supplemented with 200 ml of microbial liquid filtrate derived from washing the AM fungal inoculum mixed with soil : sand mix filtered through sieves down to 20 μm to standardise the non-AM fungal microbial community across all pots. Plants were grown in a glasshouse with day/night temperatures of 27 and 18°C, respectively, with a 12 h photoperiod. Watering was initially 100 ml every day for the first 3 wk, followed by 400–500 ml every 3 d. Volumes were adjusted as needed to maintain consistent soil moisture across pots, monitored every 3 d using a handheld moisture meter (PMS-714; Lutron Electronic Enterprise; Taipei, Taiwan). Plants were harvested 55 d from germination, where all hosts were toward the latter vegetative growth stages, before booting or flowering. For each plant, from six locations distributed across the root system, 10–15 root fragments were collected from each location. These were cut into 5-cm segments, composited and stored in 50% ethanol before mycorrhizal fungal colonisation assessment, and the remaining aboveground and belowground plant tissues were all oven-dried at 60°C. Total biomass was recorded, and all foliar tissue was ground and homogenised before any chemical analyses.
Phosphorus (P) concentrations in plants were assessed via inductively coupled plasma spectroscopy following digestions with nitric acid (Zarcinas et al., 1987). Total foliar phenolics were determined as described in Salminen & Karonen (2011), in technical triplicates, using a Folin–Ciocalteu assay with gallic acid monohydrate (Sigma-Aldrich, St Louis, MO, USA) as the quantification standard. For assessing root colonisation by AM fungi, and to confirm the absence of any root colonisation by AM fungi in the control plants, ethanol-stored root samples were placed into histology cassettes, then cleared with 10% potassium hydroxide (KOH) at 90°C for 10 min, stained with 5% ink-vinegar (using Quink ink; Parker Nantes, France) at 90°C for 15 min (Vierheilig et al., 1998). Root fragments of 5 cm each were mounted on glass slides with glycerin under a coverslip. For each plant, 30 root fragments were assessed in total, representing 150 cm of root length. The percentage of root length colonised by any AM fungal structure (total colonisation), as well as the percentage of root length colonised by arbuscules (arbuscular colonisation), and vesicles (vesicular colonisation) were examined microscopically using the intersect method at ×200 magnification for at least 100 intersections per plant (McGonigle et al., 1990). The McGonigle et al. (1990) intersect method was used as it remains the most widely applied approach (Füzy et al., 2015), allowing comparability across the literature, despite the availability of alternative methods that may offer greater sensitivity (e.g. Trouvelot, 1986; Kokkoris et al., 2019).
To examine the relationship between root length colonised (total colonisation and arbuscular colonisation) and plant outcomes, plant mycorrhizal responses were calculated as ((plant response − mean plant responses with no AM fungi)/mean plant responses with no AM fungi) × 100, where the plant response was either the total biomass, P concentration, or phenolic concentration. These provided the mycorrhizal growth responses (MGR), mycorrhizal phosphorus responses (MPR) and the mycorrhizal phenolic responses (MPhenR). Due to very low vesicular colonisation across samples, vesicles were not included in the main analyses; however, these data are provided in the Supporting Information (Fig. S1c).
Generalised additive models (GAMs) were fitted for each plant species separately. These models were initially fitted for each response variable using the mgcv package in R (Wood, 2003). Inoculation rates were used to generate variation in colonisation and were not included in GAMs as plant responses were analysed in relation to root length colonised, the biologically relevant predictor for the hypotheses. GAMs were chosen for their flexibility in capturing both linear and nonlinear trends without assuming a predefined functional form. Smooth terms were fitted using the default basis dimension (k = 10) in the mgcv package, the suitability of the k value was confirmed using diagnostic checks with the gam.check function. Model smoothness was determined using generalised cross-validation, and the effective degrees of freedom (edf) of the smooth term were examined to assess the shape of the relationship. Where the GAM indicated a significant relationship and edf was close to 1 (suggesting an approximately linear trend), a linear model (LM) was fitted and model selection was confirmed using Akaike Information Criterion (AIC). If a GAM indicated a significant relationship that was nonlinear (edf > 1.8), a nonlinear asymptotic model was tested using nonlinear least squares (NLS). The asymptotic model followed the self-starting asymptotic function SSasymp in R. Model parameters were tested for significance, and AIC values were used to confirm the best model fit. Where a GAM showed no significant relationship (P > 0.05 for the smooth term), no further modelling was conducted, and the response variable was considered to be independent of root length colonised. GAM fits were used to visualise relationships and the statistical outputs of the best fitting models (GAM, LM, or NLS) were reported on plots (Fig. 1). Predictions were obtained using the predict function over an evenly spaced sequence of colonisation values. Confidence intervals (95%) were generated from the GAM or LM and displayed as shaded regions in all plots. Where GAMs yielded non-significant relationships, no trend lines or confidence intervals were displayed in the plots. Model assumptions were checked via residual diagnostics (using the gam.check function for GAMs, Shapiro–Wilk tests for normality in LMs, and parameter stability checks for NLS models). Model selection was confirmed based on AIC comparisons and significance testing of fitted parameters. Statistical analyses were performed in R (v.4.3.3) using the mgcv (Wood, 2003), ggeffects (Lüdecke, 2018) and nlme packages (Pinheiro & Bates, 2000).
Measuring the proportion of root length colonised by AM fungi offers a logical proxy for inferring the functional outcomes of the symbiosis. Yet this study adds to others that demonstrate that the relationship between colonisation and benefit is far from straightforward. It varies across traits, species and colonisation types – and, critically, not all functional outcomes appear to scale with the proportion of root length colonised in the same way.
Among all plant responses measured, the clearest and most consistent pattern emerged in relation to plant defence chemistry. In all four plant species, the total colonisation was strongly and non-linearly associated with increases in mycorrhizal phenolic responses (MPhenR), with each exhibiting an asymptotic relationship (Table S1; Fig. 1). In all species, GAMs were significant (P ≤ 0.01), with effective degrees of freedom (edf) ranging from 1.75 in maize to 7.56 in wheat, and high explanatory power (R2 ranging from 0.32 to 0.86). These patterns were best described by NLS models supporting my second hypothesis that colonisation–response relationships would often be non-linear, and (in part) my first, which predicted positive associations between total colonisation and phenolics.
As total colonisation increased, plant phenolic responses rose sharply at low to moderate levels, before reaching a plateau. This may suggest that phenolic defence responses are triggered by the plant signalling associated with the presence of fungi within the root (Jung et al., 2012) rather than arbuscule development, and that these responses do not necessarily continue to rise in proportion to the percentage of root length colonised. That this pattern held consistently across both C3 and C4 crops did not support my first hypothesis that anticipated C4 hosts would be more responsive.
Strikingly, no significant relationships were observed between arbuscular colonisation and MPhenR in any species (GAM P > 0.05; edf ≈ 1; R2 < 0.1; Table S1; Fig. 1). While some models propose that arbuscule-mediated nutrient exchange enhances defence by enabling greater resource allocation (Pozo & Azcón-Aguilar, 2007; Vannette & Hunter, 2011), others have demonstrated that defence responses can be activated upon fungal entry, independent of arbuscule formation or nutritional benefit (Cameron et al., 2013; Anthony et al., 2020; Pozo de la Hoz et al., 2021). This decoupling suggests that phenolic responses are not mediated by the arbuscules themselves – despite their role as the primary interface for carbon and nutrient exchange. Arbuscular colonisation can sometimes have weak or inconsistent associations with plant traits due to the ephemeral nature of arbuscules, yet the consistent absence of any relationship across all hosts was unexpected, particularly when compared to the consistent relationship plant phenolic responses had with the total root length colonised. Rather than reflecting methodological noise due to inaccuracies of relying on measurements of the ephemeral arbuscules, this pattern may point to a biological disconnect: phenolic responses could be triggered by fungal presence more broadly, not by arbuscular function per se.
This distinction is important when viewed in light of the resource exchange model of plant defence from Vannette & Hunter (2011), which posited that defence benefits from AM fungi initially increase with colonisation but may diminish at higher fungal densities due to rising carbon costs and diminishing nutrient returns. By contrast, the patterns observed here – consistent defence responses even in the absence of growth or phosphorus benefits – suggest that the phenolic responses are not closely linked to carbon-for-nutrient exchange per se, as they occur even in the absence of proportional growth or phosphorus responses and are not predicted by arbuscule abundance. Rather than supporting a strict cost–benefit trade-off model, the findings here align with the understanding of a mycorrhiza-induced resistance which can be primed through early-stage fungal signalling, such as root penetration or other pre-exchange cues, independent of extensive resource exchange (Cameron et al., 2013; Anthony et al., 2020; Weinberger et al., 2025).
The saturation of the plant phenolic responses may reflect an upper threshold of defence activation, beyond which either host regulation downregulates further metabolic investment, or fungal suppression of host defence limits further accumulation. The absence of a corresponding relationship with arbuscular colonisation does challenge the idea that nutrient exchange is required for downstream defence benefits. Instead, it suggests a broader role for colonisation as a signalling event, a cue which may then trigger systemic resistance.
Together, these findings suggest that AM fungi can modulate plant phenotype in ways that are independent of resource acquisition. Smith & Smith (2011) noted that P uptake via the AM fungal pathway can occur without observable growth responses, particularly in non-responsive species or under high-P conditions. Similarly, the patterns reported here imply that AM fungi may confer consistent defence-related benefits even when classical markers of mutualism (mycorrhizal growth and P responses) show little change.
In contrast to the strong phenolic patterns, and contradicting my first hypothesis, no significant associations were observed between colonisation (total or arbuscular) and MGR across any crop species (GAM P > 0.05; R2 ≤ 0.12). The plants in my study were grown in soil with high P availability (c. 46 mg kg−1 Colwell P), which probably affected the likelihood of observing such effects. Nonetheless, this P level falls within the critical range reported for these crops (typically 22–48 mg kg−1 depending on soil type and pH), and thus the soil P here would somewhat reflect conditions typical of high-input agricultural systems in which these crops are bred and grown. Additionally, while plant root growth was largely unrestricted, minor root binding was observed in a few sorghum replicates with particularly high root biomass – an occasional artefact of pot experiments that may influence root development or function. Although limited, such constraints could have modestly contributed to the absence of stronger growth–colonisation links. Nonetheless, average MGRs were positive across all species, with sorghum exhibiting the strongest response.
Phosphorus responses (MPR) showed clearer patterns, particularly in the C4 crops. In sorghum, MPR was significantly and non-linearly associated with total colonisation (GAM edf = 1.86, R2 = 0.49, P < 0.001), and best fit by an NLS model. In maize, the relationship was linear (R2 = 0.35, P = 0.001). These findings are consistent with my first and second hypotheses and align with previous work suggesting that C4 species often derive P benefits from the symbiosis in ways more tightly linked to the proportion of root length colonised (Treseder, 2013). The asymptotic shape of the MPR response in sorghum may suggest early saturation of P benefit, while maize's linear pattern could reflect a more gradual uptake curve. These contrasting responses could challenge assumptions that P benefit may always saturate at high colonisation. Interestingly, barley showed a significant linear relationship between arbuscular colonisation and MPR (R2 = 0.41, P = 0.002), a result not predicted by my first or second hypothesis in that I expected the root length colonised in C4 plants to have stronger relationship with MPR than in C3 plants, and that this relationship would be non-linear.
Overall, the results of this study would suggest that phenolic responses may be among the most consistent outcomes of AM fungal colonisation in high-input systems, and that these may occur independently of traditional growth or nutrient-associated benefit metrics. Here I did not test whether the phenolics did confer increased resistance to pathogens or insect herbivores. This would have given a stronger basis for a functional interpretation of the MPhenR across the plant hosts.
Researchers of arbuscular mycorrhizas are well acquainted with context dependency, and the caveats here are no exception. While this study draws on data from four globally important crops, the relationships observed are likely to vary with crop variety, AM fungal identity, and environmental conditions. Rather than proposing these patterns as universal, I offer them as a prompt to revisit how we assess AM symbiosis function. This may be particularly valuable in agricultural systems, where high nutrient availability can mute growth and P mycorrhizal benefit (Ryan & Kirkegaard, 2012).
While this study aimed to interrogate the functional meaning of colonisation metrics as they are commonly used, it is clear that methodological constraints remain a barrier to deeper understanding. Traditional staining-based techniques, including the one used here, do not discriminate between living and dead fungal structures and could conflate functional colonisation with fungal necromass. Moreover, colonisation was assessed at a single endpoint, limiting insight into the temporal dynamics of the symbiosis. Recent advances, including live-imaging techniques such as AMSlides (McGaley et al., 2025) or genetically modified hosts that report arbuscule formation through anthocyanin expression (Kumar et al., 2022), offer new tools to monitor colonisation over time. Similarly, molecular approaches (Bodenhausen et al., 2021) targeting mitochondrial or nuclear markers could be used, and perhaps could help, to quantify metabolically active fungal tissue, while image-based analyses could allow more nuanced trait-based characterisation of intraradical structures, including potentially important distinctions in fungal morphology across host species. These emerging methods offer valuable opportunities to move beyond static, aggregate colonisation measures and to refine how we detect, interpret and ultimately understand the ecological and functional roles of AM fungal colonisation.
Finally, while the proportion of root-length colonised and similar measures remain useful and easily obtained metrics, I hope this Letter encourages careful interpretation. Indeed, the positive associations observed here between total colonisation and plant phenolics (in all four species), and between arbuscular colonisation and foliar phosphorus in three species, reinforce its utility as an informative, albeit context-dependent, indicator of symbiotic outcomes. Assessing colonisation remains fundamental to the study of the AM symbiosis; the presence of AM fungi inside the root is a prerequisite for the symbiosis itself. Yet, as we continue to rely on colonisation metrics to understand symbiotic function, we must ask not only what it measures, but what assumptions we carry with it. If we are to better understand the role of AM fungi in shaping plant traits and ecosystem processes, we must also sharpen how we interpret the most fundamental measures of the symbiosis.
The New Phytologist Foundation remains neutral with regard to jurisdictional claims in maps and in any institutional affiliations.
期刊介绍:
New Phytologist is an international electronic journal published 24 times a year. It is owned by the New Phytologist Foundation, a non-profit-making charitable organization dedicated to promoting plant science. The journal publishes excellent, novel, rigorous, and timely research and scholarship in plant science and its applications. The articles cover topics in five sections: Physiology & Development, Environment, Interaction, Evolution, and Transformative Plant Biotechnology. These sections encompass intracellular processes, global environmental change, and encourage cross-disciplinary approaches. The journal recognizes the use of techniques from molecular and cell biology, functional genomics, modeling, and system-based approaches in plant science. Abstracting and Indexing Information for New Phytologist includes Academic Search, AgBiotech News & Information, Agroforestry Abstracts, Biochemistry & Biophysics Citation Index, Botanical Pesticides, CAB Abstracts®, Environment Index, Global Health, and Plant Breeding Abstracts, and others.
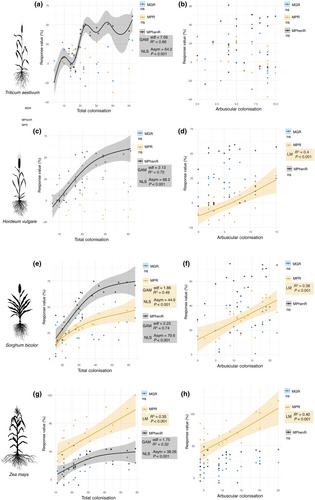

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


