Maaike G. J. M. van Bergen, Priscilla Kamphuis, Aniek O. de Graaf, Jonas B. Salzbrunn, Theresia N. Koorenhof-Scheele, Isabelle A. van Zeventer, Avinash G. Dinmohamed, Jan Jacob Schuringa, Bert A. van der Reijden, Gerwin Huls, Joop H. Jansen
{"title":"老年人群中的克隆造血和骨髓偏斜","authors":"Maaike G. J. M. van Bergen, Priscilla Kamphuis, Aniek O. de Graaf, Jonas B. Salzbrunn, Theresia N. Koorenhof-Scheele, Isabelle A. van Zeventer, Avinash G. Dinmohamed, Jan Jacob Schuringa, Bert A. van der Reijden, Gerwin Huls, Joop H. Jansen","doi":"10.1002/ajh.27495","DOIUrl":null,"url":null,"abstract":"<p>Hematopoietic stem cells (HSCs) continuously produce blood cells while maintaining their self-renewal, proliferation, and differentiation potential. Normal blood cell production is balanced between myeloid and lymphoid progeny. With aging, the number of HSCs increases but their differentiation potential declines.<span><sup>1</sup></span> One of the hallmarks of aged HSCs is a myeloid differentiation bias, with less capability of differentiation toward the lymphoid lineage, resulting in age-related myeloid skewing. Another common feature of the aging hematopoietic system is the increased prevalence of somatic driver mutations within the HSC compartment. Clonal outgrowth of a subpopulation of cells sharing a mutation in a hematological malignancy-associated driver gene is called clonal hematopoiesis (CH).<span><sup>2</sup></span> Since the prevalence of both conditions increase with age, we questioned whether there is an association between myeloid skewing and CH.</p><p>To gain insight into the changes in myeloid and lymphoid progeny upon aging, we analyzed all individuals from the Dutch population-based Lifelines cohort ≥18 years with available myeloid and lymphoid peripheral blood counts (<i>n</i> = 144 676). In males, the percentage of myeloid cells from the total leukocytes increased significantly with aging (<i>p</i> < .001; Figures S1 and S2), while in females, the myeloid cells showed a periodic pattern with an initial increase, followed by a decrease during menopause and finally increased again from the age of 70 (Figures S1 and S2). A clear difference was observed between males and females for the changes in myeloid cell counts. This may be explained by changes in sex hormone levels, as the number of neutrophils decreases significantly during menopause in females. However, we observed a clear shift in the mean percentage of myeloid cells upon aging (Figure S1).</p><p>To investigate whether there is an association between the myeloid cell percentage and CH, we evaluated all individuals ≥60 years from the Lifelines cohort (<i>n</i> = 21 727) with available myeloid and lymphoid blood cell counts from whom we had generated CH data previously (<i>n</i> = 4607; Figures S3 and S4, Supplemental Methods; Data S1, Table S1). The percentage of myeloid cells was normally distributed in this cohort with a mean of 67.8% myeloid cells (Figure S4). From these individuals, <i>n</i> = 1899 (41.2%) carried at least one driver gene mutation with a variant allele frequency (VAF) ≥1%. A significant association was observed between the percentage of myeloid cells and mutations in <i>JAK2</i> (OR 1.06, 95% CI 1.03–1.09; <i>p</i> < .001), <i>SF3B1</i> (OR 1.03, 95% CI 1.00–1.07; <i>p</i> = .034), and <i>TET2</i> (OR 1.01, 95% CI 1.00–1.02; <i>p</i> = .020; Figure S4). Overall, no significant correlation was observed between the percentage of myeloid cells and the clone size in the cohort with available myeloid cell counts and CH (<i>n</i> = 1899; <i>p</i> = .891; Figures 1A and S5). However, we observed a positive correlation between the percentage of myeloid cells and the clone size of <i>JAK2</i> (Spearman's rank correlation coefficient 0.319; <i>p</i> = .012; Figure 1B) and <i>ASXL1</i> (Spearman's rank correlation coefficient 0.279; <i>p</i> = .002; Figure 1C). In line with this, it has been shown that homozygous <i>JAK2-V617F</i> mutations associate with increased white blood cell counts compared with heterozygous mutations.<span><sup>3</sup></span> Our data suggest that this dosage effect may already be present in a premalignant heterozygous state.</p><p>Subsequently, we investigated the association between CH and aberrant myeloid cell counts upon aging. In the absence of a generally accepted, fixed threshold to describe myeloid skewing upon aging, we selected individuals ≥60 years with the highest (99th) percentile (<i>n</i> = 218, 138 males, 80 females, myeloid percentage ≥ 83.82%) and lowest (1st) percentile (<i>n</i> = 218, 47 males, 171 females, myeloid percentage ≤ 46.08%) of myeloid cells, as well as 1:1 age- and sex-matched controls (Tables S2 and S3; Figures S6 and S7). The prevalence of CH was not significantly different in high myeloid cases compared with their matched controls (42.8% vs. 38.9%; <i>p</i> = .431; Figure S7), and low myeloid cases compared with their matched controls (33.3% vs. 36.4%; <i>p</i> = .543). Despite their low frequency, we observed significantly more mutations in spliceosome-associated genes (<i>SF3B1</i> and <i>SRSF2</i>) compared with their matched controls (4.2% vs. 0.5%; <i>p</i> = .020; Figures 1D,G, S8, and S9). All spliceosome mutations were detected in cases with concurrent anemia (<i>n</i> = 9). We subsequently investigated the clone size in cases with low or high myeloid cell percentages and observed a significantly larger clone size in <i>TET2</i> mutant low myeloid cases compared with their matched controls (median VAF 13.0% vs. 2.0%; <i>p</i> = .005; Figure 1H), while the clone sizes of high myeloid cases were not significantly different from controls (Figure 1E). We hypothesize that low myeloid cases carrying <i>TET2</i> mutations represent cases with an underlying lymphoid malignancy, as <i>TET2</i> mutations are commonly identified in, for example, lymphoma or diffuse large B-cell lymphoma.<span><sup>4</sup></span></p><p>To determine whether CH-associated mutations are restricted to cells derived from the myeloid or lymphoid lineage in high or low myeloid cases, we sorted mature myeloid and lymphoid cells from selected cases and established the presence and clone size of the mutations (Supplemental Methods; Data S1). We selected samples that carried mutations in <i>DNMT3A</i> or <i>TET2</i>, as these are the most commonly mutated genes. Overall, the VAF was increased in cells derived from the myeloid lineage (granulocytes, monocytes) compared with cells derived from the lymphoid lineage (T- and B-lymphocytes; Figure S10). The VAF of the mutated NK-cell clones were comparable to the myeloid lineage (monocytes, granulocytes).<span><sup>5</sup></span> Although we observed a significant increase in the <i>TET2</i> mutant clone size of low myeloid cases compared with their matched controls, no specific enrichment of <i>TET2</i> mutations in the lymphoid fraction compared with the myeloid fraction was revealed (Tables S4 and S5).</p><p>Follow-up data were available from a subset of cases with a high or low myeloid percentage but showed no significant difference in the proportion of CH between cases with a consistently high or low myeloid percentage at follow-up or cases who corrected their myeloid cell percentage (Figures S11 and S12). Furthermore, the changes of myeloid cell percentages over time did not depend on the presence of CH at baseline (Figure S11) nor the clone size (Figure S13). Cases with a high myeloid percentage had a significantly higher number of platelets (<i>p</i> = .023) and the marker for inflammation, high-sensitive C-reactive protein (hsCRP), was significantly increased in cases with a high myeloid cell percentage compared with the matched controls (<i>p</i> < .001, Table S2), while in cases with a low myeloid percentage the platelet counts (<i>p</i> = .035) and hsCRP levels (<i>p</i> = .015) were significantly decreased. The elevated hsCRP level could be a contributing factor to myeloid skewing.</p><p>To evaluate the consequences of aberrant myeloid cell counts, we determined whether the presence of CH impacted the survival of cases with high or low myeloid skewing. High myeloid skewing cases have inferior survival compared with their age- and sex-matched controls (<i>p</i> < .001; HR 2.47, 95% CI 1.54–3.97), which is in line with studies showing that a high Neutrophil-to-Lymphocyte Ratio (NLR) is associated with all-cause mortality.<span><sup>6</sup></span> However, the presence of CH did not significantly affect survival of high myeloid skewing cases (<i>p</i> = .427; HR 1.25, 95% CI 0.72–2.19; Figure 1F; Table S6A). Furthermore, low myeloid skewing cases showed a trend toward inferior survival compared with their matched controls (<i>p</i> = .051; HR 2.19, 95% CI 1.00–4.82), but the presence of CH did not have an effect on survival in these cases (<i>p</i> = .345; HR 1.53, 95% CI 0.63–3.72; Figure 1I; Table S6B). When stratifying the analysis for specific mutated genes, cases with low myeloid skewing showed a higher all-cause mortality for individuals carrying <i>TET2</i> mutations (<i>p</i> = .020; HR 3.42, 95% CI 1.22–9.64; Figure S4; Table S7).</p><p>By establishing linkage of the Lifelines cohort to the nationwide population-based National Cancer Registry database, we investigated the incident diagnoses of hematological neoplasms. First, we evaluated the cumulative incidence of hematological malignancies in all individuals ≥60 years from the Lifelines cohort with available myeloid cell counts (<i>n</i> = 21 599) with correction for age and sex. A higher cumulative incidence was observed in individuals with the highest myeloid cell percentage (>80%, <i>n</i> = 739; <i>p</i> < .001; HR 3.13, 95% CI 1.94–5.03) and lowest myeloid cell percentage (≤50%, <i>n</i> = 621; <i>p</i> < .001; HR 3.06, 95% CI 1.76–5.32; Figure S14). Furthermore, a higher cumulative incidence was observed in individuals with a myeloid cell percentage of 51–60% (<i>p</i> = .011; HR 1.55, 95% CI 1.11–2.17; Figure S14). Thereafter, we evaluated the cumulative incidence of hematological malignancies in high or low myeloid cases compared with their matched controls. Due to the low number of incidences, we could not stratify our analysis for the presence of CH. After a follow-up period of 10 years, 6 high myeloid cases were diagnosed with hematological malignancies (2.8% of the cases, <i>n</i> = 3 developed myeloid malignancies and <i>n</i> = 3 developed lymphoid malignancies), which showed a higher cumulative incidence compare with their matched controls, although this did not reach statistical significance (<i>p</i> = .052; Figure 1J). The incidence of hematological malignancies was comparable among low myeloid cases (<i>n</i> = 6 incident diagnoses, 2.9%, <i>n</i> = 2 developed myeloid malignancies and <i>n</i> = 4 developed lymphoid malignancies) and their matched controls (<i>p</i> = .316; Figure 1K). Together, we show that the absolute number as well as the relative abundance of myeloid over lymphoid cells increases significantly upon aging, but that this is not driven by the presence of clonal hematopoiesis (CH).</p><p>M.G.J.M.v.B., P.K., A.O.d.G., J.B.S., T.N.K., and I.A.v.Z. contributed to study design, data collection, analysis, and interpretation of the data; J.J.S., A.G.D., and B.A.v.d.R. were involved in the interpretation of the data; G.H. and J.H.J. were principal investigators and involved in the study design, data collection, and interpretation of the results; M.G.J.M.v.B and P.K. wrote the manuscript that was critically revised by all co-authors.</p><p>This work was supported by the MDS-RIGHT project, which has received funding from the European Union's Horizon 2020 research and innovation program under grant agreement 634789. The funder of this study had no role in study design; collection, analysis, and interpretation of data; and writing or approval of the manuscript. The Lifelines Biobank initiative has been made possible by subsidy from the Dutch Ministry of Health, Welfare and Sport; the Dutch Ministry of Economic Affairs; the University Medical Center Groningen; University Groningen; and the Northern Provinces of The Netherlands. A.O.d.G., M.G.J.M.v.B., and J.H.J. were supported by a grant from the Dutch Cancer Society (grant number 10813).</p><p>The authors declare no conflicts of interest.</p>","PeriodicalId":7724,"journal":{"name":"American Journal of Hematology","volume":"99 12","pages":"2402-2405"},"PeriodicalIF":10.1000,"publicationDate":"2024-10-21","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1002/ajh.27495","citationCount":"0","resultStr":"{\"title\":\"Clonal hematopoiesis and myeloid skewing in older population-based individuals\",\"authors\":\"Maaike G. J. M. van Bergen, Priscilla Kamphuis, Aniek O. de Graaf, Jonas B. Salzbrunn, Theresia N. Koorenhof-Scheele, Isabelle A. van Zeventer, Avinash G. Dinmohamed, Jan Jacob Schuringa, Bert A. van der Reijden, Gerwin Huls, Joop H. Jansen\",\"doi\":\"10.1002/ajh.27495\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p>Hematopoietic stem cells (HSCs) continuously produce blood cells while maintaining their self-renewal, proliferation, and differentiation potential. Normal blood cell production is balanced between myeloid and lymphoid progeny. With aging, the number of HSCs increases but their differentiation potential declines.<span><sup>1</sup></span> One of the hallmarks of aged HSCs is a myeloid differentiation bias, with less capability of differentiation toward the lymphoid lineage, resulting in age-related myeloid skewing. Another common feature of the aging hematopoietic system is the increased prevalence of somatic driver mutations within the HSC compartment. Clonal outgrowth of a subpopulation of cells sharing a mutation in a hematological malignancy-associated driver gene is called clonal hematopoiesis (CH).<span><sup>2</sup></span> Since the prevalence of both conditions increase with age, we questioned whether there is an association between myeloid skewing and CH.</p><p>To gain insight into the changes in myeloid and lymphoid progeny upon aging, we analyzed all individuals from the Dutch population-based Lifelines cohort ≥18 years with available myeloid and lymphoid peripheral blood counts (<i>n</i> = 144 676). In males, the percentage of myeloid cells from the total leukocytes increased significantly with aging (<i>p</i> < .001; Figures S1 and S2), while in females, the myeloid cells showed a periodic pattern with an initial increase, followed by a decrease during menopause and finally increased again from the age of 70 (Figures S1 and S2). A clear difference was observed between males and females for the changes in myeloid cell counts. This may be explained by changes in sex hormone levels, as the number of neutrophils decreases significantly during menopause in females. However, we observed a clear shift in the mean percentage of myeloid cells upon aging (Figure S1).</p><p>To investigate whether there is an association between the myeloid cell percentage and CH, we evaluated all individuals ≥60 years from the Lifelines cohort (<i>n</i> = 21 727) with available myeloid and lymphoid blood cell counts from whom we had generated CH data previously (<i>n</i> = 4607; Figures S3 and S4, Supplemental Methods; Data S1, Table S1). The percentage of myeloid cells was normally distributed in this cohort with a mean of 67.8% myeloid cells (Figure S4). From these individuals, <i>n</i> = 1899 (41.2%) carried at least one driver gene mutation with a variant allele frequency (VAF) ≥1%. A significant association was observed between the percentage of myeloid cells and mutations in <i>JAK2</i> (OR 1.06, 95% CI 1.03–1.09; <i>p</i> < .001), <i>SF3B1</i> (OR 1.03, 95% CI 1.00–1.07; <i>p</i> = .034), and <i>TET2</i> (OR 1.01, 95% CI 1.00–1.02; <i>p</i> = .020; Figure S4). Overall, no significant correlation was observed between the percentage of myeloid cells and the clone size in the cohort with available myeloid cell counts and CH (<i>n</i> = 1899; <i>p</i> = .891; Figures 1A and S5). However, we observed a positive correlation between the percentage of myeloid cells and the clone size of <i>JAK2</i> (Spearman's rank correlation coefficient 0.319; <i>p</i> = .012; Figure 1B) and <i>ASXL1</i> (Spearman's rank correlation coefficient 0.279; <i>p</i> = .002; Figure 1C). In line with this, it has been shown that homozygous <i>JAK2-V617F</i> mutations associate with increased white blood cell counts compared with heterozygous mutations.<span><sup>3</sup></span> Our data suggest that this dosage effect may already be present in a premalignant heterozygous state.</p><p>Subsequently, we investigated the association between CH and aberrant myeloid cell counts upon aging. In the absence of a generally accepted, fixed threshold to describe myeloid skewing upon aging, we selected individuals ≥60 years with the highest (99th) percentile (<i>n</i> = 218, 138 males, 80 females, myeloid percentage ≥ 83.82%) and lowest (1st) percentile (<i>n</i> = 218, 47 males, 171 females, myeloid percentage ≤ 46.08%) of myeloid cells, as well as 1:1 age- and sex-matched controls (Tables S2 and S3; Figures S6 and S7). The prevalence of CH was not significantly different in high myeloid cases compared with their matched controls (42.8% vs. 38.9%; <i>p</i> = .431; Figure S7), and low myeloid cases compared with their matched controls (33.3% vs. 36.4%; <i>p</i> = .543). Despite their low frequency, we observed significantly more mutations in spliceosome-associated genes (<i>SF3B1</i> and <i>SRSF2</i>) compared with their matched controls (4.2% vs. 0.5%; <i>p</i> = .020; Figures 1D,G, S8, and S9). All spliceosome mutations were detected in cases with concurrent anemia (<i>n</i> = 9). We subsequently investigated the clone size in cases with low or high myeloid cell percentages and observed a significantly larger clone size in <i>TET2</i> mutant low myeloid cases compared with their matched controls (median VAF 13.0% vs. 2.0%; <i>p</i> = .005; Figure 1H), while the clone sizes of high myeloid cases were not significantly different from controls (Figure 1E). We hypothesize that low myeloid cases carrying <i>TET2</i> mutations represent cases with an underlying lymphoid malignancy, as <i>TET2</i> mutations are commonly identified in, for example, lymphoma or diffuse large B-cell lymphoma.<span><sup>4</sup></span></p><p>To determine whether CH-associated mutations are restricted to cells derived from the myeloid or lymphoid lineage in high or low myeloid cases, we sorted mature myeloid and lymphoid cells from selected cases and established the presence and clone size of the mutations (Supplemental Methods; Data S1). We selected samples that carried mutations in <i>DNMT3A</i> or <i>TET2</i>, as these are the most commonly mutated genes. Overall, the VAF was increased in cells derived from the myeloid lineage (granulocytes, monocytes) compared with cells derived from the lymphoid lineage (T- and B-lymphocytes; Figure S10). The VAF of the mutated NK-cell clones were comparable to the myeloid lineage (monocytes, granulocytes).<span><sup>5</sup></span> Although we observed a significant increase in the <i>TET2</i> mutant clone size of low myeloid cases compared with their matched controls, no specific enrichment of <i>TET2</i> mutations in the lymphoid fraction compared with the myeloid fraction was revealed (Tables S4 and S5).</p><p>Follow-up data were available from a subset of cases with a high or low myeloid percentage but showed no significant difference in the proportion of CH between cases with a consistently high or low myeloid percentage at follow-up or cases who corrected their myeloid cell percentage (Figures S11 and S12). Furthermore, the changes of myeloid cell percentages over time did not depend on the presence of CH at baseline (Figure S11) nor the clone size (Figure S13). Cases with a high myeloid percentage had a significantly higher number of platelets (<i>p</i> = .023) and the marker for inflammation, high-sensitive C-reactive protein (hsCRP), was significantly increased in cases with a high myeloid cell percentage compared with the matched controls (<i>p</i> < .001, Table S2), while in cases with a low myeloid percentage the platelet counts (<i>p</i> = .035) and hsCRP levels (<i>p</i> = .015) were significantly decreased. The elevated hsCRP level could be a contributing factor to myeloid skewing.</p><p>To evaluate the consequences of aberrant myeloid cell counts, we determined whether the presence of CH impacted the survival of cases with high or low myeloid skewing. High myeloid skewing cases have inferior survival compared with their age- and sex-matched controls (<i>p</i> < .001; HR 2.47, 95% CI 1.54–3.97), which is in line with studies showing that a high Neutrophil-to-Lymphocyte Ratio (NLR) is associated with all-cause mortality.<span><sup>6</sup></span> However, the presence of CH did not significantly affect survival of high myeloid skewing cases (<i>p</i> = .427; HR 1.25, 95% CI 0.72–2.19; Figure 1F; Table S6A). Furthermore, low myeloid skewing cases showed a trend toward inferior survival compared with their matched controls (<i>p</i> = .051; HR 2.19, 95% CI 1.00–4.82), but the presence of CH did not have an effect on survival in these cases (<i>p</i> = .345; HR 1.53, 95% CI 0.63–3.72; Figure 1I; Table S6B). When stratifying the analysis for specific mutated genes, cases with low myeloid skewing showed a higher all-cause mortality for individuals carrying <i>TET2</i> mutations (<i>p</i> = .020; HR 3.42, 95% CI 1.22–9.64; Figure S4; Table S7).</p><p>By establishing linkage of the Lifelines cohort to the nationwide population-based National Cancer Registry database, we investigated the incident diagnoses of hematological neoplasms. First, we evaluated the cumulative incidence of hematological malignancies in all individuals ≥60 years from the Lifelines cohort with available myeloid cell counts (<i>n</i> = 21 599) with correction for age and sex. A higher cumulative incidence was observed in individuals with the highest myeloid cell percentage (>80%, <i>n</i> = 739; <i>p</i> < .001; HR 3.13, 95% CI 1.94–5.03) and lowest myeloid cell percentage (≤50%, <i>n</i> = 621; <i>p</i> < .001; HR 3.06, 95% CI 1.76–5.32; Figure S14). Furthermore, a higher cumulative incidence was observed in individuals with a myeloid cell percentage of 51–60% (<i>p</i> = .011; HR 1.55, 95% CI 1.11–2.17; Figure S14). Thereafter, we evaluated the cumulative incidence of hematological malignancies in high or low myeloid cases compared with their matched controls. Due to the low number of incidences, we could not stratify our analysis for the presence of CH. After a follow-up period of 10 years, 6 high myeloid cases were diagnosed with hematological malignancies (2.8% of the cases, <i>n</i> = 3 developed myeloid malignancies and <i>n</i> = 3 developed lymphoid malignancies), which showed a higher cumulative incidence compare with their matched controls, although this did not reach statistical significance (<i>p</i> = .052; Figure 1J). The incidence of hematological malignancies was comparable among low myeloid cases (<i>n</i> = 6 incident diagnoses, 2.9%, <i>n</i> = 2 developed myeloid malignancies and <i>n</i> = 4 developed lymphoid malignancies) and their matched controls (<i>p</i> = .316; Figure 1K). Together, we show that the absolute number as well as the relative abundance of myeloid over lymphoid cells increases significantly upon aging, but that this is not driven by the presence of clonal hematopoiesis (CH).</p><p>M.G.J.M.v.B., P.K., A.O.d.G., J.B.S., T.N.K., and I.A.v.Z. contributed to study design, data collection, analysis, and interpretation of the data; J.J.S., A.G.D., and B.A.v.d.R. were involved in the interpretation of the data; G.H. and J.H.J. were principal investigators and involved in the study design, data collection, and interpretation of the results; M.G.J.M.v.B and P.K. wrote the manuscript that was critically revised by all co-authors.</p><p>This work was supported by the MDS-RIGHT project, which has received funding from the European Union's Horizon 2020 research and innovation program under grant agreement 634789. The funder of this study had no role in study design; collection, analysis, and interpretation of data; and writing or approval of the manuscript. The Lifelines Biobank initiative has been made possible by subsidy from the Dutch Ministry of Health, Welfare and Sport; the Dutch Ministry of Economic Affairs; the University Medical Center Groningen; University Groningen; and the Northern Provinces of The Netherlands. A.O.d.G., M.G.J.M.v.B., and J.H.J. were supported by a grant from the Dutch Cancer Society (grant number 10813).</p><p>The authors declare no conflicts of interest.</p>\",\"PeriodicalId\":7724,\"journal\":{\"name\":\"American Journal of Hematology\",\"volume\":\"99 12\",\"pages\":\"2402-2405\"},\"PeriodicalIF\":10.1000,\"publicationDate\":\"2024-10-21\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://onlinelibrary.wiley.com/doi/epdf/10.1002/ajh.27495\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"American Journal of Hematology\",\"FirstCategoryId\":\"3\",\"ListUrlMain\":\"https://onlinelibrary.wiley.com/doi/10.1002/ajh.27495\",\"RegionNum\":1,\"RegionCategory\":\"医学\",\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"Q1\",\"JCRName\":\"HEMATOLOGY\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"American Journal of Hematology","FirstCategoryId":"3","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1002/ajh.27495","RegionNum":1,"RegionCategory":"医学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"HEMATOLOGY","Score":null,"Total":0}
Clonal hematopoiesis and myeloid skewing in older population-based individuals
Hematopoietic stem cells (HSCs) continuously produce blood cells while maintaining their self-renewal, proliferation, and differentiation potential. Normal blood cell production is balanced between myeloid and lymphoid progeny. With aging, the number of HSCs increases but their differentiation potential declines.1 One of the hallmarks of aged HSCs is a myeloid differentiation bias, with less capability of differentiation toward the lymphoid lineage, resulting in age-related myeloid skewing. Another common feature of the aging hematopoietic system is the increased prevalence of somatic driver mutations within the HSC compartment. Clonal outgrowth of a subpopulation of cells sharing a mutation in a hematological malignancy-associated driver gene is called clonal hematopoiesis (CH).2 Since the prevalence of both conditions increase with age, we questioned whether there is an association between myeloid skewing and CH.
To gain insight into the changes in myeloid and lymphoid progeny upon aging, we analyzed all individuals from the Dutch population-based Lifelines cohort ≥18 years with available myeloid and lymphoid peripheral blood counts (n = 144 676). In males, the percentage of myeloid cells from the total leukocytes increased significantly with aging (p < .001; Figures S1 and S2), while in females, the myeloid cells showed a periodic pattern with an initial increase, followed by a decrease during menopause and finally increased again from the age of 70 (Figures S1 and S2). A clear difference was observed between males and females for the changes in myeloid cell counts. This may be explained by changes in sex hormone levels, as the number of neutrophils decreases significantly during menopause in females. However, we observed a clear shift in the mean percentage of myeloid cells upon aging (Figure S1).
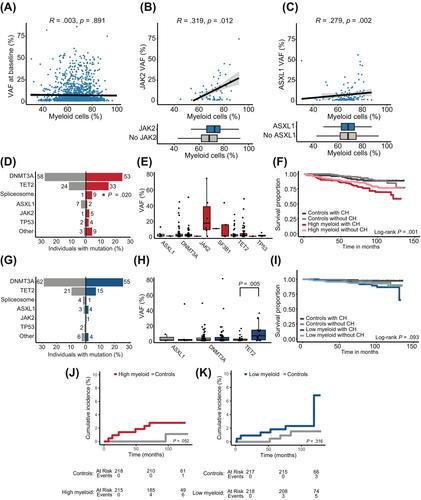
To investigate whether there is an association between the myeloid cell percentage and CH, we evaluated all individuals ≥60 years from the Lifelines cohort (n = 21 727) with available myeloid and lymphoid blood cell counts from whom we had generated CH data previously (n = 4607; Figures S3 and S4, Supplemental Methods; Data S1, Table S1). The percentage of myeloid cells was normally distributed in this cohort with a mean of 67.8% myeloid cells (Figure S4). From these individuals, n = 1899 (41.2%) carried at least one driver gene mutation with a variant allele frequency (VAF) ≥1%. A significant association was observed between the percentage of myeloid cells and mutations in JAK2 (OR 1.06, 95% CI 1.03–1.09; p < .001), SF3B1 (OR 1.03, 95% CI 1.00–1.07; p = .034), and TET2 (OR 1.01, 95% CI 1.00–1.02; p = .020; Figure S4). Overall, no significant correlation was observed between the percentage of myeloid cells and the clone size in the cohort with available myeloid cell counts and CH (n = 1899; p = .891; Figures 1A and S5). However, we observed a positive correlation between the percentage of myeloid cells and the clone size of JAK2 (Spearman's rank correlation coefficient 0.319; p = .012; Figure 1B) and ASXL1 (Spearman's rank correlation coefficient 0.279; p = .002; Figure 1C). In line with this, it has been shown that homozygous JAK2-V617F mutations associate with increased white blood cell counts compared with heterozygous mutations.3 Our data suggest that this dosage effect may already be present in a premalignant heterozygous state.
Subsequently, we investigated the association between CH and aberrant myeloid cell counts upon aging. In the absence of a generally accepted, fixed threshold to describe myeloid skewing upon aging, we selected individuals ≥60 years with the highest (99th) percentile (n = 218, 138 males, 80 females, myeloid percentage ≥ 83.82%) and lowest (1st) percentile (n = 218, 47 males, 171 females, myeloid percentage ≤ 46.08%) of myeloid cells, as well as 1:1 age- and sex-matched controls (Tables S2 and S3; Figures S6 and S7). The prevalence of CH was not significantly different in high myeloid cases compared with their matched controls (42.8% vs. 38.9%; p = .431; Figure S7), and low myeloid cases compared with their matched controls (33.3% vs. 36.4%; p = .543). Despite their low frequency, we observed significantly more mutations in spliceosome-associated genes (SF3B1 and SRSF2) compared with their matched controls (4.2% vs. 0.5%; p = .020; Figures 1D,G, S8, and S9). All spliceosome mutations were detected in cases with concurrent anemia (n = 9). We subsequently investigated the clone size in cases with low or high myeloid cell percentages and observed a significantly larger clone size in TET2 mutant low myeloid cases compared with their matched controls (median VAF 13.0% vs. 2.0%; p = .005; Figure 1H), while the clone sizes of high myeloid cases were not significantly different from controls (Figure 1E). We hypothesize that low myeloid cases carrying TET2 mutations represent cases with an underlying lymphoid malignancy, as TET2 mutations are commonly identified in, for example, lymphoma or diffuse large B-cell lymphoma.4
To determine whether CH-associated mutations are restricted to cells derived from the myeloid or lymphoid lineage in high or low myeloid cases, we sorted mature myeloid and lymphoid cells from selected cases and established the presence and clone size of the mutations (Supplemental Methods; Data S1). We selected samples that carried mutations in DNMT3A or TET2, as these are the most commonly mutated genes. Overall, the VAF was increased in cells derived from the myeloid lineage (granulocytes, monocytes) compared with cells derived from the lymphoid lineage (T- and B-lymphocytes; Figure S10). The VAF of the mutated NK-cell clones were comparable to the myeloid lineage (monocytes, granulocytes).5 Although we observed a significant increase in the TET2 mutant clone size of low myeloid cases compared with their matched controls, no specific enrichment of TET2 mutations in the lymphoid fraction compared with the myeloid fraction was revealed (Tables S4 and S5).
Follow-up data were available from a subset of cases with a high or low myeloid percentage but showed no significant difference in the proportion of CH between cases with a consistently high or low myeloid percentage at follow-up or cases who corrected their myeloid cell percentage (Figures S11 and S12). Furthermore, the changes of myeloid cell percentages over time did not depend on the presence of CH at baseline (Figure S11) nor the clone size (Figure S13). Cases with a high myeloid percentage had a significantly higher number of platelets (p = .023) and the marker for inflammation, high-sensitive C-reactive protein (hsCRP), was significantly increased in cases with a high myeloid cell percentage compared with the matched controls (p < .001, Table S2), while in cases with a low myeloid percentage the platelet counts (p = .035) and hsCRP levels (p = .015) were significantly decreased. The elevated hsCRP level could be a contributing factor to myeloid skewing.
To evaluate the consequences of aberrant myeloid cell counts, we determined whether the presence of CH impacted the survival of cases with high or low myeloid skewing. High myeloid skewing cases have inferior survival compared with their age- and sex-matched controls (p < .001; HR 2.47, 95% CI 1.54–3.97), which is in line with studies showing that a high Neutrophil-to-Lymphocyte Ratio (NLR) is associated with all-cause mortality.6 However, the presence of CH did not significantly affect survival of high myeloid skewing cases (p = .427; HR 1.25, 95% CI 0.72–2.19; Figure 1F; Table S6A). Furthermore, low myeloid skewing cases showed a trend toward inferior survival compared with their matched controls (p = .051; HR 2.19, 95% CI 1.00–4.82), but the presence of CH did not have an effect on survival in these cases (p = .345; HR 1.53, 95% CI 0.63–3.72; Figure 1I; Table S6B). When stratifying the analysis for specific mutated genes, cases with low myeloid skewing showed a higher all-cause mortality for individuals carrying TET2 mutations (p = .020; HR 3.42, 95% CI 1.22–9.64; Figure S4; Table S7).
By establishing linkage of the Lifelines cohort to the nationwide population-based National Cancer Registry database, we investigated the incident diagnoses of hematological neoplasms. First, we evaluated the cumulative incidence of hematological malignancies in all individuals ≥60 years from the Lifelines cohort with available myeloid cell counts (n = 21 599) with correction for age and sex. A higher cumulative incidence was observed in individuals with the highest myeloid cell percentage (>80%, n = 739; p < .001; HR 3.13, 95% CI 1.94–5.03) and lowest myeloid cell percentage (≤50%, n = 621; p < .001; HR 3.06, 95% CI 1.76–5.32; Figure S14). Furthermore, a higher cumulative incidence was observed in individuals with a myeloid cell percentage of 51–60% (p = .011; HR 1.55, 95% CI 1.11–2.17; Figure S14). Thereafter, we evaluated the cumulative incidence of hematological malignancies in high or low myeloid cases compared with their matched controls. Due to the low number of incidences, we could not stratify our analysis for the presence of CH. After a follow-up period of 10 years, 6 high myeloid cases were diagnosed with hematological malignancies (2.8% of the cases, n = 3 developed myeloid malignancies and n = 3 developed lymphoid malignancies), which showed a higher cumulative incidence compare with their matched controls, although this did not reach statistical significance (p = .052; Figure 1J). The incidence of hematological malignancies was comparable among low myeloid cases (n = 6 incident diagnoses, 2.9%, n = 2 developed myeloid malignancies and n = 4 developed lymphoid malignancies) and their matched controls (p = .316; Figure 1K). Together, we show that the absolute number as well as the relative abundance of myeloid over lymphoid cells increases significantly upon aging, but that this is not driven by the presence of clonal hematopoiesis (CH).
M.G.J.M.v.B., P.K., A.O.d.G., J.B.S., T.N.K., and I.A.v.Z. contributed to study design, data collection, analysis, and interpretation of the data; J.J.S., A.G.D., and B.A.v.d.R. were involved in the interpretation of the data; G.H. and J.H.J. were principal investigators and involved in the study design, data collection, and interpretation of the results; M.G.J.M.v.B and P.K. wrote the manuscript that was critically revised by all co-authors.
This work was supported by the MDS-RIGHT project, which has received funding from the European Union's Horizon 2020 research and innovation program under grant agreement 634789. The funder of this study had no role in study design; collection, analysis, and interpretation of data; and writing or approval of the manuscript. The Lifelines Biobank initiative has been made possible by subsidy from the Dutch Ministry of Health, Welfare and Sport; the Dutch Ministry of Economic Affairs; the University Medical Center Groningen; University Groningen; and the Northern Provinces of The Netherlands. A.O.d.G., M.G.J.M.v.B., and J.H.J. were supported by a grant from the Dutch Cancer Society (grant number 10813).
期刊介绍:
The American Journal of Hematology offers extensive coverage of experimental and clinical aspects of blood diseases in humans and animal models. The journal publishes original contributions in both non-malignant and malignant hematological diseases, encompassing clinical and basic studies in areas such as hemostasis, thrombosis, immunology, blood banking, and stem cell biology. Clinical translational reports highlighting innovative therapeutic approaches for the diagnosis and treatment of hematological diseases are actively encouraged.The American Journal of Hematology features regular original laboratory and clinical research articles, brief research reports, critical reviews, images in hematology, as well as letters and correspondence.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


