棉花的植株间防御诱导是通过草食性植物侵害时挥发物的延迟释放来实现的
IF 8.3
1区 生物学
Q1 PLANT SCIENCES
引用次数: 0
摘要
引言 植物会产生多种次级代谢产物,使其能够抵御食草动物和病原体等敌害。这些化合物可以作为毒素,直接降低食草动物的存活率或繁殖成功率(如醌类、生物碱、花青素和萜类化合物),或者作为间接防御信号,如挥发性有机化合物(VOCs)(Pichersky & Lewinsohn, 2011; Mithöfer & Boland, 2012; Kessler & Kalske, 2018; Pichersky & Raguso, 2018)。这些挥发性有机化合物可以储存并持续释放(Gershenzon,1994 年,2000 年;Clancy 等人,2016 年),也可以在被食草动物捕食后被诱导并从头合成(Paré & Tumlinson,1997 年)。重要的是,这些食草动物诱导的变化包括植物释放的挥发性混合物的成分和相对比例的变化(Turlings & Erb, 2018),其中包含与生态相关的攻击风险提示。食草动物诱导的植物挥发物(HIPVs)可以驱赶食草动物并吸引其敌人;它们还可以作为单株植物不同部分之间的信号(植物内部信号),以激活预防性系统防御(Heil & Silva Bueno, 2007; Meents & Mithöfer, 2020),并可能被邻近植物用来为未来的攻击做好准备(Morrell & Kessler, 2017; Schuman, 2023)。最初的发现表明,植物之间通过挥发性介导的相互作用来应对食草动物的攻击(Baldwin & Schultz, 1983; Farmer & Ryan, 1990; Bruin et al、1992)受到了一些怀疑,但现在已被广泛接受,认为其既常见又与生态相关(Heil & Karban, 2010; Ninkovic 等人,2019; Kessler 等人,2023)。许多研究报告了由 HIPVs 介导的植物间信号传递的作用(Baldwin & Schultz, 1983; Dolch & Tscharntke, 2000; Karban 等人,2003; Heil & Silva Bueno, 2007),实地研究揭示了所涉及的挥发性线索的特异性(Karban 等人,2004; Moreira 等人,2016; Kalske 等人,2019)。据报道,作为潜在信号线索的食草动物诱导植物挥发物包括茉莉酸盐(Farmer & Ryan, 1990)、绿叶挥发物(Engelberth & Engelberth, 2019)和芳香化合物(Erb 等人,2015)。这些来自受损植物的 HIPV 可以到达未受损的邻近植物,从而进入所谓的 "引诱 "状态(Ton 等人,2007 年;Mauch-Mani 等人,2017 年)。虽然被激活的植物有时不表达防御或只表达低水平的防御,但这些植物在受到攻击后会表现出大大增强的防御化合物诱导(Conrath 等人,2006 年;Martinez-Medina 等人,2016 年)。此外,暴露于 HIPVs 的未受损植物也会立即提高防御能力,而无需与攻击者直接接触;这些诱导的防御能力会在食草动物攻击之前就已存在(Karban 等人,2003 年;Waterman 等人,2024 年)。棉花虫害严重,需要大量施用杀虫剂,占全球杀虫剂用量的很大一部分(Coupe & Capel, 2016; Huang et al.)虽然这些化学品的使用提高了作物产量,但也对环境造成了极为不利的影响(Van Der Werf,1996 年;Aktar 等人,2009 年),尤其是土壤和水污染。人们正在寻求更加良性的害虫控制策略,包括增强植物的自然防御能力(Llandres 等人,2018 年)。众所周知,格桑花可通过定量和定性地改变其挥发性排放特征,以及增加其非挥发性防御性萜类醛的含量(如格桑酚和氦化物)来应对昆虫的食草行为(Loughrin 等人,1994 年;McCall 等人,1994 年;Röse 等人,1996 年;McAuslane 等人,1997 年;Arce 等人,2021 年)。有趣的是,受损植物释放出的挥发性混合物也会随着草食动物攻击开始后的时间而变化,受损后立即释放出储存的挥发性化合物(如萜烯 α-蒎烯和叶绿素),而新合成的化合物则会在攻击开始后至少 24 小时后大量释放出来(Loughrin 等人,1994 年;Paré & Tumlinson, 1997 年)。后一种化合物包括萜烯类化合物,如 β-ocimene 和 β-farnesene 以及芳香吲哚,这些化合物在未受损或刚受损的植物中释放量极低或根本不释放。众所周知,在实验室和田间条件下,受到食草螨攻击的棉花植株比未受损植株更能抵抗螨类的新定植(Karban,1985 年,1986 年)。本文章由计算机程序翻译,如有差异,请以英文原文为准。
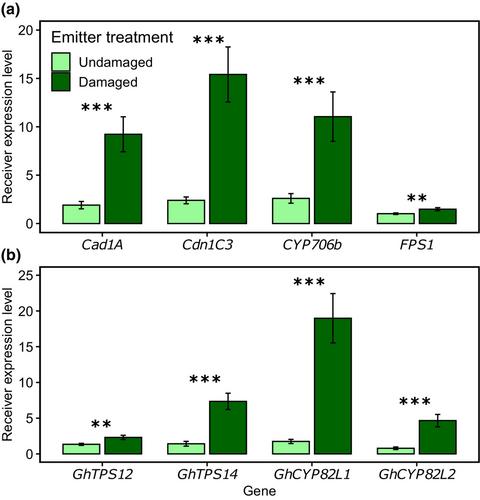
Plant-to-plant defence induction in cotton is mediated by delayed release of volatiles upon herbivory
求助全文
通过发布文献求助,成功后即可免费获取论文全文。
去求助
来源期刊

New Phytologist
生物-植物科学
自引率
5.30%
发文量
728
期刊介绍:
New Phytologist is an international electronic journal published 24 times a year. It is owned by the New Phytologist Foundation, a non-profit-making charitable organization dedicated to promoting plant science. The journal publishes excellent, novel, rigorous, and timely research and scholarship in plant science and its applications. The articles cover topics in five sections: Physiology & Development, Environment, Interaction, Evolution, and Transformative Plant Biotechnology. These sections encompass intracellular processes, global environmental change, and encourage cross-disciplinary approaches. The journal recognizes the use of techniques from molecular and cell biology, functional genomics, modeling, and system-based approaches in plant science. Abstracting and Indexing Information for New Phytologist includes Academic Search, AgBiotech News & Information, Agroforestry Abstracts, Biochemistry & Biophysics Citation Index, Botanical Pesticides, CAB Abstracts®, Environment Index, Global Health, and Plant Breeding Abstracts, and others.
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


