{"title":"植物-微生物-微生物关联的化学语言:虚拟议题导言。","authors":"Stéphane Hacquard, Francis M. Martin","doi":"10.1111/nph.20124","DOIUrl":null,"url":null,"abstract":"<p>The chemical language between plants and microbes, also known as interspecies chemical communication, is a sophisticated system of signal exchange involving a diverse array of molecular compounds that regulate and mediate complex host–microbe interactions and drive high-level biological organization. This intricate communication network encompasses primary and specialized metabolites that underpin host–microbe nutrient exchange, host–microbe assembly processes, or plant–soil feedbacks that ultimately explain host–microbiota associations, as well as plant health and disease states. Some of these metabolites (such as phytohormones, specialized metabolites, volatile organic compounds, and peptides) can act as signaling molecules, which plants and microbes produce, perceive, and respond to, thereby facilitating symbiotic relationships, pathogen defense, and environmental adaptation. Co-evolution between plants and microbiota members, as well as between microbiota members that show stable associations with plants over evolutionary time, is a critical aspect of their chemical communication strategies, where co-adapted organisms undergo reciprocal evolutionary changes selecting or counter-selecting for specific associations. This dynamic process is expected to shape both host and microbial genomes, behaviors, and ecological roles, leading to interdependent and sometimes highly specialized relationships explaining the diversity, specificity, and stability of plant–microbiota interactions. This dynamic and complex chemical dialogue is also predicted to be modulated by environmental factors and specific biological contexts, reflecting eco-evolutionary adaptations that ultimately influence ecosystem functions and stability.</p><p>In this Virtual Issue, we aim to showcase <i>New Phytologist</i>'s commitment to plant microbiome research by highlighting recent articles and reviews that aim to unravel the chemical language of plant–microbe–microbe associations. Experts in this field explore open questions and future research lines, including: <i>How do plant exudates shape the phylogenetic diversity and physiology of plant microbiota? Which host or microbial metabolites shape microbiota establishment or drive host-specific signatures in microbiota assemblies across plant species? Which microbial and host metabolites/antimicrobials protect against pests and pathogens and how can they be used to promote plant health in agriculture? Are specialized plant metabolites involved in more complex feedback loops with microbiota members that drive host phenotypes and/or stress adaptation? What are the current open questions, research needs and priorities?</i> This Virtual Issue illustrates that the chemical language between plants and microbes, as well as among microbes, is not only critical for understanding high-level biological organization and beneficial plant–microbiota associations, but also a prerequisite for advancing agricultural sustainability and innovation (Fig. 1).</p><p>Root-exuded metabolites play a critical role in altering belowground microbiota establishment, influencing plant–soil interactions, and promoting microbe-induced stress resistance (Hong <i>et al</i>., <span>2022</span>). A previous report revealed remarkable similarity in microbiota composition between the model grass <i>Brachypodium distachyon</i> and the biocrust cyanobacterium <i>Microcoleus vaginatus</i>. Therefore, the core microbiome reproducibly found in association with phylogenetically diverse photosynthetic organisms is likely explained by evolutionarily conserved photosynthesis-derived molecules or byproducts (Zheng <i>et al</i>., <span>2023</span>). By contrast, host-specific specialized metabolites have emerged as key determinants that drive species-specific signatures in plant-associated microbiota. For example, in Brassicaceae, glucosinolates represent lineage-specific metabolic innovations with defensive functions against herbivores and microbial pathogens. A recent report indicated that endoplasmic reticulum (ER) bodies and their resident myrosinases (enzymes that hydrolyze glucosinolates) can modulate the profile of root-secreted indole glucosinolates in <i>Arabidopsis</i>, thereby influencing the microbiota assembly at the root interface (Basak <i>et al</i>., <span>2024</span>). Similarly, natural rubber biosynthesis was also found to reduce herbivory and to modulate plant–microbiota establishment, illustrating the importance and multiple roles of plant-specialized metabolites in shaping multitrophic interactions (Böttner <i>et al</i>., <span>2023</span>). Compounds exuded by photosynthetic organisms can selectively promote the growth of specific microbiota members. For instance, algal exudates influence the composition of the surrounding bacterial community through specific algal metabolites that act as selective bacterial substrates (such as 4-hydroxybenzoic acid), and thus represents a mechanism by which algal exudates modulate microbiota assembly (Brisson <i>et al</i>., <span>2023</span>). Host-derived metabolites not only selectively shape the composition but also the function of microbiota members. This is illustrated by the observation that 16 root-exuded metabolites were found to significantly regulate functional gene expression in plant growth-promoting rhizobacteria (Feng <i>et al</i>., <span>2023</span>). Taken together, these data support the idea that host-derived metabolites are key determinants of the assembly and function of the plant microbiota (Fig. 1).</p><p>Plant–soil feedbacks represent chemical, biological and physical legacies that plants leave in the soil, and which can positively or negatively influence the growth of other plants. In maize, a class of tryptophan-derived metabolites abundantly found in root exudates (called benzoxazinoids) was found to increase maize resistance to negative plant–soil feedback when grown in soils conditioned by winter wheat, lacy phacelia, and rapeseed (Gfeller <i>et al</i>., <span>2024</span>). Notably, soil sterilization and microbiota reinoculation experiments suggest that this benzoxazinoid-mediated protection acts via changes in the composition and/or function of the soil microbiota. Consistent with this, another report showed that soil treatment with an allelochemical (i.e. a chemical released from a donor that affects the growth of a receiver) found in rosemary scrub soils alters the soil microbiota composition and revealed that allelopathic inhibition of plant productivity is mitigated, at least to some extent, by the microbiota (Revillini <i>et al</i>., <span>2023</span>). Allelopathy-mediated plant growth inhibition occurs not only between distantly related plant species but also, more surprisingly, within the same plant species. For example, in poplar, female roots release a greater amount and more diverse phenolic allelochemicals into the soil environment, resulting in growth inhibition of same-sex neighbors. Compared with female monocultures, the presence of males reduced total phenolic accumulation in the soil, resulting in a shift from allelopathic inhibition to chemical facilitation, which was also surprisingly microbiota-dependent (Xia <i>et al</i>., <span>2023</span>). Finally, another report indicated that the benefits of intercropping on plant growth and nutrient uptake were largely attributed to the interactions between rhizosphere metabolites and microbiomes. Indeed, the exogenous application of key metabolites enriched by intercropping significantly promoted maize biomass in natural soil, but not in sterilized soil (Jiang <i>et al</i>., <span>2024</span>). Taken together, these data highlight the potential of chemical-selected microbiomes to modulate plant health and illustrate that both positive and negative plant–soil feedbacks observed in nature are, at least to some extent, microbiota-dependent (Fig. 1).</p><p>Microbes can influence the production of secondary metabolites and hormones in plants, which are crucial for plant defense, signaling, and interactions with other organisms or with the environment. In the Amaryllidaceae plant <i>Lycoris radiata</i>, bacteria insensitive to Amaryllidaceae alkaloids (AAs) have been shown to stimulate the accumulation of these alkaloids in leaves (Zhou <i>et al</i>., <span>2024</span>). This creates a positive feedback loop that promotes the establishment of AA-insensitive strains over AA-sensitive strains and resistance to a fungal pathogen. In a different study, it was reported that distinct soil microbiomes can independently influence the flavor chemistry of mustard seeds (<i>Brassica juncea</i>) by altering the glucosinolate content (Walsh <i>et al</i>., <span>2024</span>). The authors identified specific rhizosphere microbial taxa and bacterial genes related to sulfur metabolism that predicted the concentration of the main glucosinolate, allyl, in seeds. This work highlights the potential role of belowground microbiomes not only in promoting defense chemical accumulation in seeds but also in altering the flavor profiles of seeds (Nakano, <span>2024</span>; Walsh <i>et al</i>., <span>2024</span>). Another report indicated that a keystone bacterial taxon, which is consistently enriched in the seed microbiome (Simonin <i>et al</i>., <span>2022</span>), is essential for the evolution of herbicide resistance in weeds (Hu <i>et al</i>., <span>2024</span>; Zhang <i>et al</i>., <span>2024</span>). Hu <i>et al</i>. (<span>2024</span>) revealed that this microbiota-induced herbicide resistance is more likely explained by modulation of host xenobiotic metabolism and betalain biosynthesis pathways than by the bacterial capability to directly catabolize the herbicide. Chemically selected microbiota members have also been found to induce a reactive oxygen species (ROS) burst in cucumber roots, thereby contributing to disease suppression (Wen <i>et al</i>., <span>2023</span>). Finally, another report by Zhang <i>et al</i>. (<span>2023</span>) revealed that the soil-borne bacterium <i>Klebsiella pneumoniae</i> can induce cluster root formation in white lupin via the modulation of ethylene signaling, leading to enhanced host nutrient uptake. The authors proposed that high 1-aminocyclopropane-1-carboxylate deaminase activity in <i>Klebsiella aerogenes</i> inhibits the biosynthesis of ethylene in plants by cleaving the ethylene precursor 1-aminocyclopropane-1-carboxylic acid, thus promoting cluster root formation under phosphate-limiting conditions (Zhang <i>et al</i>., <span>2023</span>). These examples illustrate how microbes can alter the host chemical landscape and underpin the relevance of microbiota-induced changes in host chemistry for plant productivity (Fig. 1).</p><p>Members of the microbiota inhibit pathogens through mechanisms such as chemical production and nutrient competition. They produce antimicrobial compounds that directly inhibit or kill pathogens, and secrete enzymes that degrade pathogen structures. For example, analysis of age-related microbiota in <i>Populus tomentosa</i> shows how mature trees develop more diverse microbial communities that are gradually enriched in Actinobacterial taxa (Xie <i>et al</i>., <span>2023</span>). Corresponding isolates efficiently suppress tree diseases through direct antimicrobial-mediated pathogen suppression and therefore likely supplement host innate immunity to protect older trees in nature. Enrichment of host-protective taxa has also been observed in the context of crop rotation. Hong <i>et al</i>. (<span>2023</span>) revealed that crop rotation-induced suppression of banana wilt disease was associated with rotation-enriched microbial taxa that displayed strong antagonistic activity against <i>Fusarium oxysporum</i> pathogens and protected the host from disease. Additionally, microbiota members compete with pathogens for essential nutrients and space, effectively limiting access to pathogens and growth opportunities. For example, Zhou <i>et al</i>. (<span>2023</span>) tested the influence of carbon resource diversity on the microbial trophic network architecture and pathogen invasion success using tomato microcosm experiments. The authors concluded that expanding the carbon source diversity increased microbial evenness, widened the microbial functional landscape, and promoted disease suppression via efficient resource competition with pathogens (Zhou <i>et al</i>., <span>2023</span>). Similarly, the critical function of host-derived sugars for bacterial niche competition in flowers and for the modulation of virulence factor expression in pathogens has recently been discussed by Lee <i>et al</i>. (<span>2024</span>). These authors observed that flower-derived arabinogalactan is an important sugar that drives pathogen colonization, and that competition for arabinogalactan between the pathogen and the sympatric commensal <i>Pantoea agglomerans</i> is likely key for <i>P. agglomerans</i>-mediated disease suppression. Furthermore, arabinogalactan consumption by <i>P. agglomerans</i> alters the production of major virulence determinants in the pathogen (Lee <i>et al</i>., <span>2024</span>). These findings underscore the importance of microbe-mediated chemical and nutrient blocking for successful niche colonization and pathogen suppression (Fig. 1).</p><p>Over time, host-adapted pathogens have evolved strategies to manipulate microbiota to promote infection. Several reports present compelling evidence that filamentous plant pathogens can manipulate plant microbiota to their advantage by targeting beneficial microbial communities. Although pathogens are also known to alter microbiota assembly through chemical and nutrient competition, several reports published in this Virtual Issue indicate that they also use effector proteins and enzymes to do so. For instance, the <i>Verticillium dahliae</i> effector gene <i>VdAve1L</i> exhibits extensive allelic variation. Snelders <i>et al</i>. (<span>2023</span>) demonstrated that the full-length effector variant <i>VdAve1L2</i> has antimicrobial activity, which the fungus uses to mediate tomato colonization by directly suppressing the antagonistic Actinobacteria in the host microbiota. Similarly, the parasitic protist <i>Albugo candida</i> releases proteins into the apoplast of the plant that selectively suppress beneficial phyllosphere-associated bacteria (gram-positive) (Gómez-Pérez <i>et al</i>., <span>2023</span>; Rovenich & Thomma, <span>2023</span>). These authors observed that intrinsically disordered regions of the protein and a high-positive net charge are often associated with the antibacterial activity of the proteins (Gómez-Pérez <i>et al</i>., <span>2023</span>). Finally, another report revealed that the smut fungus <i>Ustilago maydis</i> exclusively expresses a gene encoding ribotoxin (Ribo1) at early infection stages (Ökmen <i>et al</i>., <span>2023</span>). This secreted ribonuclease has a wide range of cytotoxic activities against bacteria and yeast cells and enables smut fungi to compete with host-associated bacteria. Taken together, these strategies enable pathogens to outcompete host-associated bacteria to enhance their colonization (Fig. 1).</p><p>In conclusion, we hope that this Virtual Issue nicely illustrates that chemical communication between organisms drives high-level biological organization and ecosystem functioning, and represents a key mechanism by which plants and microbes adapt to their often-stressful surrounding environments (Fig. 1). The remarkable diversity of molecules and mechanisms deployed by living organisms to respond to environmental cues is key for species survival and therefore contributes to landscape diversity.</p>","PeriodicalId":214,"journal":{"name":"New Phytologist","volume":"244 3","pages":"739-742"},"PeriodicalIF":8.3000,"publicationDate":"2024-10-02","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1111/nph.20124","citationCount":"0","resultStr":"{\"title\":\"The chemical language of plant–microbe–microbe associations: an introduction to a Virtual Issue\",\"authors\":\"Stéphane Hacquard, Francis M. Martin\",\"doi\":\"10.1111/nph.20124\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p>The chemical language between plants and microbes, also known as interspecies chemical communication, is a sophisticated system of signal exchange involving a diverse array of molecular compounds that regulate and mediate complex host–microbe interactions and drive high-level biological organization. This intricate communication network encompasses primary and specialized metabolites that underpin host–microbe nutrient exchange, host–microbe assembly processes, or plant–soil feedbacks that ultimately explain host–microbiota associations, as well as plant health and disease states. Some of these metabolites (such as phytohormones, specialized metabolites, volatile organic compounds, and peptides) can act as signaling molecules, which plants and microbes produce, perceive, and respond to, thereby facilitating symbiotic relationships, pathogen defense, and environmental adaptation. Co-evolution between plants and microbiota members, as well as between microbiota members that show stable associations with plants over evolutionary time, is a critical aspect of their chemical communication strategies, where co-adapted organisms undergo reciprocal evolutionary changes selecting or counter-selecting for specific associations. This dynamic process is expected to shape both host and microbial genomes, behaviors, and ecological roles, leading to interdependent and sometimes highly specialized relationships explaining the diversity, specificity, and stability of plant–microbiota interactions. This dynamic and complex chemical dialogue is also predicted to be modulated by environmental factors and specific biological contexts, reflecting eco-evolutionary adaptations that ultimately influence ecosystem functions and stability.</p><p>In this Virtual Issue, we aim to showcase <i>New Phytologist</i>'s commitment to plant microbiome research by highlighting recent articles and reviews that aim to unravel the chemical language of plant–microbe–microbe associations. Experts in this field explore open questions and future research lines, including: <i>How do plant exudates shape the phylogenetic diversity and physiology of plant microbiota? Which host or microbial metabolites shape microbiota establishment or drive host-specific signatures in microbiota assemblies across plant species? Which microbial and host metabolites/antimicrobials protect against pests and pathogens and how can they be used to promote plant health in agriculture? Are specialized plant metabolites involved in more complex feedback loops with microbiota members that drive host phenotypes and/or stress adaptation? What are the current open questions, research needs and priorities?</i> This Virtual Issue illustrates that the chemical language between plants and microbes, as well as among microbes, is not only critical for understanding high-level biological organization and beneficial plant–microbiota associations, but also a prerequisite for advancing agricultural sustainability and innovation (Fig. 1).</p><p>Root-exuded metabolites play a critical role in altering belowground microbiota establishment, influencing plant–soil interactions, and promoting microbe-induced stress resistance (Hong <i>et al</i>., <span>2022</span>). A previous report revealed remarkable similarity in microbiota composition between the model grass <i>Brachypodium distachyon</i> and the biocrust cyanobacterium <i>Microcoleus vaginatus</i>. Therefore, the core microbiome reproducibly found in association with phylogenetically diverse photosynthetic organisms is likely explained by evolutionarily conserved photosynthesis-derived molecules or byproducts (Zheng <i>et al</i>., <span>2023</span>). By contrast, host-specific specialized metabolites have emerged as key determinants that drive species-specific signatures in plant-associated microbiota. For example, in Brassicaceae, glucosinolates represent lineage-specific metabolic innovations with defensive functions against herbivores and microbial pathogens. A recent report indicated that endoplasmic reticulum (ER) bodies and their resident myrosinases (enzymes that hydrolyze glucosinolates) can modulate the profile of root-secreted indole glucosinolates in <i>Arabidopsis</i>, thereby influencing the microbiota assembly at the root interface (Basak <i>et al</i>., <span>2024</span>). Similarly, natural rubber biosynthesis was also found to reduce herbivory and to modulate plant–microbiota establishment, illustrating the importance and multiple roles of plant-specialized metabolites in shaping multitrophic interactions (Böttner <i>et al</i>., <span>2023</span>). Compounds exuded by photosynthetic organisms can selectively promote the growth of specific microbiota members. For instance, algal exudates influence the composition of the surrounding bacterial community through specific algal metabolites that act as selective bacterial substrates (such as 4-hydroxybenzoic acid), and thus represents a mechanism by which algal exudates modulate microbiota assembly (Brisson <i>et al</i>., <span>2023</span>). Host-derived metabolites not only selectively shape the composition but also the function of microbiota members. This is illustrated by the observation that 16 root-exuded metabolites were found to significantly regulate functional gene expression in plant growth-promoting rhizobacteria (Feng <i>et al</i>., <span>2023</span>). Taken together, these data support the idea that host-derived metabolites are key determinants of the assembly and function of the plant microbiota (Fig. 1).</p><p>Plant–soil feedbacks represent chemical, biological and physical legacies that plants leave in the soil, and which can positively or negatively influence the growth of other plants. In maize, a class of tryptophan-derived metabolites abundantly found in root exudates (called benzoxazinoids) was found to increase maize resistance to negative plant–soil feedback when grown in soils conditioned by winter wheat, lacy phacelia, and rapeseed (Gfeller <i>et al</i>., <span>2024</span>). Notably, soil sterilization and microbiota reinoculation experiments suggest that this benzoxazinoid-mediated protection acts via changes in the composition and/or function of the soil microbiota. Consistent with this, another report showed that soil treatment with an allelochemical (i.e. a chemical released from a donor that affects the growth of a receiver) found in rosemary scrub soils alters the soil microbiota composition and revealed that allelopathic inhibition of plant productivity is mitigated, at least to some extent, by the microbiota (Revillini <i>et al</i>., <span>2023</span>). Allelopathy-mediated plant growth inhibition occurs not only between distantly related plant species but also, more surprisingly, within the same plant species. For example, in poplar, female roots release a greater amount and more diverse phenolic allelochemicals into the soil environment, resulting in growth inhibition of same-sex neighbors. Compared with female monocultures, the presence of males reduced total phenolic accumulation in the soil, resulting in a shift from allelopathic inhibition to chemical facilitation, which was also surprisingly microbiota-dependent (Xia <i>et al</i>., <span>2023</span>). Finally, another report indicated that the benefits of intercropping on plant growth and nutrient uptake were largely attributed to the interactions between rhizosphere metabolites and microbiomes. Indeed, the exogenous application of key metabolites enriched by intercropping significantly promoted maize biomass in natural soil, but not in sterilized soil (Jiang <i>et al</i>., <span>2024</span>). Taken together, these data highlight the potential of chemical-selected microbiomes to modulate plant health and illustrate that both positive and negative plant–soil feedbacks observed in nature are, at least to some extent, microbiota-dependent (Fig. 1).</p><p>Microbes can influence the production of secondary metabolites and hormones in plants, which are crucial for plant defense, signaling, and interactions with other organisms or with the environment. In the Amaryllidaceae plant <i>Lycoris radiata</i>, bacteria insensitive to Amaryllidaceae alkaloids (AAs) have been shown to stimulate the accumulation of these alkaloids in leaves (Zhou <i>et al</i>., <span>2024</span>). This creates a positive feedback loop that promotes the establishment of AA-insensitive strains over AA-sensitive strains and resistance to a fungal pathogen. In a different study, it was reported that distinct soil microbiomes can independently influence the flavor chemistry of mustard seeds (<i>Brassica juncea</i>) by altering the glucosinolate content (Walsh <i>et al</i>., <span>2024</span>). The authors identified specific rhizosphere microbial taxa and bacterial genes related to sulfur metabolism that predicted the concentration of the main glucosinolate, allyl, in seeds. This work highlights the potential role of belowground microbiomes not only in promoting defense chemical accumulation in seeds but also in altering the flavor profiles of seeds (Nakano, <span>2024</span>; Walsh <i>et al</i>., <span>2024</span>). Another report indicated that a keystone bacterial taxon, which is consistently enriched in the seed microbiome (Simonin <i>et al</i>., <span>2022</span>), is essential for the evolution of herbicide resistance in weeds (Hu <i>et al</i>., <span>2024</span>; Zhang <i>et al</i>., <span>2024</span>). Hu <i>et al</i>. (<span>2024</span>) revealed that this microbiota-induced herbicide resistance is more likely explained by modulation of host xenobiotic metabolism and betalain biosynthesis pathways than by the bacterial capability to directly catabolize the herbicide. Chemically selected microbiota members have also been found to induce a reactive oxygen species (ROS) burst in cucumber roots, thereby contributing to disease suppression (Wen <i>et al</i>., <span>2023</span>). Finally, another report by Zhang <i>et al</i>. (<span>2023</span>) revealed that the soil-borne bacterium <i>Klebsiella pneumoniae</i> can induce cluster root formation in white lupin via the modulation of ethylene signaling, leading to enhanced host nutrient uptake. The authors proposed that high 1-aminocyclopropane-1-carboxylate deaminase activity in <i>Klebsiella aerogenes</i> inhibits the biosynthesis of ethylene in plants by cleaving the ethylene precursor 1-aminocyclopropane-1-carboxylic acid, thus promoting cluster root formation under phosphate-limiting conditions (Zhang <i>et al</i>., <span>2023</span>). These examples illustrate how microbes can alter the host chemical landscape and underpin the relevance of microbiota-induced changes in host chemistry for plant productivity (Fig. 1).</p><p>Members of the microbiota inhibit pathogens through mechanisms such as chemical production and nutrient competition. They produce antimicrobial compounds that directly inhibit or kill pathogens, and secrete enzymes that degrade pathogen structures. For example, analysis of age-related microbiota in <i>Populus tomentosa</i> shows how mature trees develop more diverse microbial communities that are gradually enriched in Actinobacterial taxa (Xie <i>et al</i>., <span>2023</span>). Corresponding isolates efficiently suppress tree diseases through direct antimicrobial-mediated pathogen suppression and therefore likely supplement host innate immunity to protect older trees in nature. Enrichment of host-protective taxa has also been observed in the context of crop rotation. Hong <i>et al</i>. (<span>2023</span>) revealed that crop rotation-induced suppression of banana wilt disease was associated with rotation-enriched microbial taxa that displayed strong antagonistic activity against <i>Fusarium oxysporum</i> pathogens and protected the host from disease. Additionally, microbiota members compete with pathogens for essential nutrients and space, effectively limiting access to pathogens and growth opportunities. For example, Zhou <i>et al</i>. (<span>2023</span>) tested the influence of carbon resource diversity on the microbial trophic network architecture and pathogen invasion success using tomato microcosm experiments. The authors concluded that expanding the carbon source diversity increased microbial evenness, widened the microbial functional landscape, and promoted disease suppression via efficient resource competition with pathogens (Zhou <i>et al</i>., <span>2023</span>). Similarly, the critical function of host-derived sugars for bacterial niche competition in flowers and for the modulation of virulence factor expression in pathogens has recently been discussed by Lee <i>et al</i>. (<span>2024</span>). These authors observed that flower-derived arabinogalactan is an important sugar that drives pathogen colonization, and that competition for arabinogalactan between the pathogen and the sympatric commensal <i>Pantoea agglomerans</i> is likely key for <i>P. agglomerans</i>-mediated disease suppression. Furthermore, arabinogalactan consumption by <i>P. agglomerans</i> alters the production of major virulence determinants in the pathogen (Lee <i>et al</i>., <span>2024</span>). These findings underscore the importance of microbe-mediated chemical and nutrient blocking for successful niche colonization and pathogen suppression (Fig. 1).</p><p>Over time, host-adapted pathogens have evolved strategies to manipulate microbiota to promote infection. Several reports present compelling evidence that filamentous plant pathogens can manipulate plant microbiota to their advantage by targeting beneficial microbial communities. Although pathogens are also known to alter microbiota assembly through chemical and nutrient competition, several reports published in this Virtual Issue indicate that they also use effector proteins and enzymes to do so. For instance, the <i>Verticillium dahliae</i> effector gene <i>VdAve1L</i> exhibits extensive allelic variation. Snelders <i>et al</i>. (<span>2023</span>) demonstrated that the full-length effector variant <i>VdAve1L2</i> has antimicrobial activity, which the fungus uses to mediate tomato colonization by directly suppressing the antagonistic Actinobacteria in the host microbiota. Similarly, the parasitic protist <i>Albugo candida</i> releases proteins into the apoplast of the plant that selectively suppress beneficial phyllosphere-associated bacteria (gram-positive) (Gómez-Pérez <i>et al</i>., <span>2023</span>; Rovenich & Thomma, <span>2023</span>). These authors observed that intrinsically disordered regions of the protein and a high-positive net charge are often associated with the antibacterial activity of the proteins (Gómez-Pérez <i>et al</i>., <span>2023</span>). Finally, another report revealed that the smut fungus <i>Ustilago maydis</i> exclusively expresses a gene encoding ribotoxin (Ribo1) at early infection stages (Ökmen <i>et al</i>., <span>2023</span>). This secreted ribonuclease has a wide range of cytotoxic activities against bacteria and yeast cells and enables smut fungi to compete with host-associated bacteria. Taken together, these strategies enable pathogens to outcompete host-associated bacteria to enhance their colonization (Fig. 1).</p><p>In conclusion, we hope that this Virtual Issue nicely illustrates that chemical communication between organisms drives high-level biological organization and ecosystem functioning, and represents a key mechanism by which plants and microbes adapt to their often-stressful surrounding environments (Fig. 1). The remarkable diversity of molecules and mechanisms deployed by living organisms to respond to environmental cues is key for species survival and therefore contributes to landscape diversity.</p>\",\"PeriodicalId\":214,\"journal\":{\"name\":\"New Phytologist\",\"volume\":\"244 3\",\"pages\":\"739-742\"},\"PeriodicalIF\":8.3000,\"publicationDate\":\"2024-10-02\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://onlinelibrary.wiley.com/doi/epdf/10.1111/nph.20124\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"New Phytologist\",\"FirstCategoryId\":\"99\",\"ListUrlMain\":\"https://onlinelibrary.wiley.com/doi/10.1111/nph.20124\",\"RegionNum\":1,\"RegionCategory\":\"生物学\",\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"Q1\",\"JCRName\":\"PLANT SCIENCES\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"New Phytologist","FirstCategoryId":"99","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1111/nph.20124","RegionNum":1,"RegionCategory":"生物学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"PLANT SCIENCES","Score":null,"Total":0}
The chemical language of plant–microbe–microbe associations: an introduction to a Virtual Issue
The chemical language between plants and microbes, also known as interspecies chemical communication, is a sophisticated system of signal exchange involving a diverse array of molecular compounds that regulate and mediate complex host–microbe interactions and drive high-level biological organization. This intricate communication network encompasses primary and specialized metabolites that underpin host–microbe nutrient exchange, host–microbe assembly processes, or plant–soil feedbacks that ultimately explain host–microbiota associations, as well as plant health and disease states. Some of these metabolites (such as phytohormones, specialized metabolites, volatile organic compounds, and peptides) can act as signaling molecules, which plants and microbes produce, perceive, and respond to, thereby facilitating symbiotic relationships, pathogen defense, and environmental adaptation. Co-evolution between plants and microbiota members, as well as between microbiota members that show stable associations with plants over evolutionary time, is a critical aspect of their chemical communication strategies, where co-adapted organisms undergo reciprocal evolutionary changes selecting or counter-selecting for specific associations. This dynamic process is expected to shape both host and microbial genomes, behaviors, and ecological roles, leading to interdependent and sometimes highly specialized relationships explaining the diversity, specificity, and stability of plant–microbiota interactions. This dynamic and complex chemical dialogue is also predicted to be modulated by environmental factors and specific biological contexts, reflecting eco-evolutionary adaptations that ultimately influence ecosystem functions and stability.
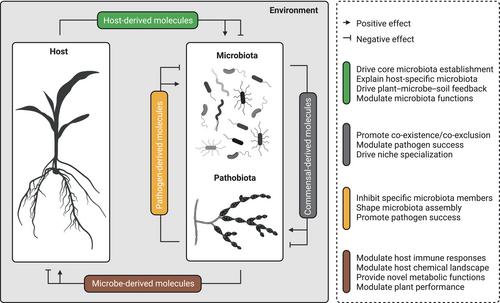
In this Virtual Issue, we aim to showcase New Phytologist's commitment to plant microbiome research by highlighting recent articles and reviews that aim to unravel the chemical language of plant–microbe–microbe associations. Experts in this field explore open questions and future research lines, including: How do plant exudates shape the phylogenetic diversity and physiology of plant microbiota? Which host or microbial metabolites shape microbiota establishment or drive host-specific signatures in microbiota assemblies across plant species? Which microbial and host metabolites/antimicrobials protect against pests and pathogens and how can they be used to promote plant health in agriculture? Are specialized plant metabolites involved in more complex feedback loops with microbiota members that drive host phenotypes and/or stress adaptation? What are the current open questions, research needs and priorities? This Virtual Issue illustrates that the chemical language between plants and microbes, as well as among microbes, is not only critical for understanding high-level biological organization and beneficial plant–microbiota associations, but also a prerequisite for advancing agricultural sustainability and innovation (Fig. 1).
Root-exuded metabolites play a critical role in altering belowground microbiota establishment, influencing plant–soil interactions, and promoting microbe-induced stress resistance (Hong et al., 2022). A previous report revealed remarkable similarity in microbiota composition between the model grass Brachypodium distachyon and the biocrust cyanobacterium Microcoleus vaginatus. Therefore, the core microbiome reproducibly found in association with phylogenetically diverse photosynthetic organisms is likely explained by evolutionarily conserved photosynthesis-derived molecules or byproducts (Zheng et al., 2023). By contrast, host-specific specialized metabolites have emerged as key determinants that drive species-specific signatures in plant-associated microbiota. For example, in Brassicaceae, glucosinolates represent lineage-specific metabolic innovations with defensive functions against herbivores and microbial pathogens. A recent report indicated that endoplasmic reticulum (ER) bodies and their resident myrosinases (enzymes that hydrolyze glucosinolates) can modulate the profile of root-secreted indole glucosinolates in Arabidopsis, thereby influencing the microbiota assembly at the root interface (Basak et al., 2024). Similarly, natural rubber biosynthesis was also found to reduce herbivory and to modulate plant–microbiota establishment, illustrating the importance and multiple roles of plant-specialized metabolites in shaping multitrophic interactions (Böttner et al., 2023). Compounds exuded by photosynthetic organisms can selectively promote the growth of specific microbiota members. For instance, algal exudates influence the composition of the surrounding bacterial community through specific algal metabolites that act as selective bacterial substrates (such as 4-hydroxybenzoic acid), and thus represents a mechanism by which algal exudates modulate microbiota assembly (Brisson et al., 2023). Host-derived metabolites not only selectively shape the composition but also the function of microbiota members. This is illustrated by the observation that 16 root-exuded metabolites were found to significantly regulate functional gene expression in plant growth-promoting rhizobacteria (Feng et al., 2023). Taken together, these data support the idea that host-derived metabolites are key determinants of the assembly and function of the plant microbiota (Fig. 1).
Plant–soil feedbacks represent chemical, biological and physical legacies that plants leave in the soil, and which can positively or negatively influence the growth of other plants. In maize, a class of tryptophan-derived metabolites abundantly found in root exudates (called benzoxazinoids) was found to increase maize resistance to negative plant–soil feedback when grown in soils conditioned by winter wheat, lacy phacelia, and rapeseed (Gfeller et al., 2024). Notably, soil sterilization and microbiota reinoculation experiments suggest that this benzoxazinoid-mediated protection acts via changes in the composition and/or function of the soil microbiota. Consistent with this, another report showed that soil treatment with an allelochemical (i.e. a chemical released from a donor that affects the growth of a receiver) found in rosemary scrub soils alters the soil microbiota composition and revealed that allelopathic inhibition of plant productivity is mitigated, at least to some extent, by the microbiota (Revillini et al., 2023). Allelopathy-mediated plant growth inhibition occurs not only between distantly related plant species but also, more surprisingly, within the same plant species. For example, in poplar, female roots release a greater amount and more diverse phenolic allelochemicals into the soil environment, resulting in growth inhibition of same-sex neighbors. Compared with female monocultures, the presence of males reduced total phenolic accumulation in the soil, resulting in a shift from allelopathic inhibition to chemical facilitation, which was also surprisingly microbiota-dependent (Xia et al., 2023). Finally, another report indicated that the benefits of intercropping on plant growth and nutrient uptake were largely attributed to the interactions between rhizosphere metabolites and microbiomes. Indeed, the exogenous application of key metabolites enriched by intercropping significantly promoted maize biomass in natural soil, but not in sterilized soil (Jiang et al., 2024). Taken together, these data highlight the potential of chemical-selected microbiomes to modulate plant health and illustrate that both positive and negative plant–soil feedbacks observed in nature are, at least to some extent, microbiota-dependent (Fig. 1).
Microbes can influence the production of secondary metabolites and hormones in plants, which are crucial for plant defense, signaling, and interactions with other organisms or with the environment. In the Amaryllidaceae plant Lycoris radiata, bacteria insensitive to Amaryllidaceae alkaloids (AAs) have been shown to stimulate the accumulation of these alkaloids in leaves (Zhou et al., 2024). This creates a positive feedback loop that promotes the establishment of AA-insensitive strains over AA-sensitive strains and resistance to a fungal pathogen. In a different study, it was reported that distinct soil microbiomes can independently influence the flavor chemistry of mustard seeds (Brassica juncea) by altering the glucosinolate content (Walsh et al., 2024). The authors identified specific rhizosphere microbial taxa and bacterial genes related to sulfur metabolism that predicted the concentration of the main glucosinolate, allyl, in seeds. This work highlights the potential role of belowground microbiomes not only in promoting defense chemical accumulation in seeds but also in altering the flavor profiles of seeds (Nakano, 2024; Walsh et al., 2024). Another report indicated that a keystone bacterial taxon, which is consistently enriched in the seed microbiome (Simonin et al., 2022), is essential for the evolution of herbicide resistance in weeds (Hu et al., 2024; Zhang et al., 2024). Hu et al. (2024) revealed that this microbiota-induced herbicide resistance is more likely explained by modulation of host xenobiotic metabolism and betalain biosynthesis pathways than by the bacterial capability to directly catabolize the herbicide. Chemically selected microbiota members have also been found to induce a reactive oxygen species (ROS) burst in cucumber roots, thereby contributing to disease suppression (Wen et al., 2023). Finally, another report by Zhang et al. (2023) revealed that the soil-borne bacterium Klebsiella pneumoniae can induce cluster root formation in white lupin via the modulation of ethylene signaling, leading to enhanced host nutrient uptake. The authors proposed that high 1-aminocyclopropane-1-carboxylate deaminase activity in Klebsiella aerogenes inhibits the biosynthesis of ethylene in plants by cleaving the ethylene precursor 1-aminocyclopropane-1-carboxylic acid, thus promoting cluster root formation under phosphate-limiting conditions (Zhang et al., 2023). These examples illustrate how microbes can alter the host chemical landscape and underpin the relevance of microbiota-induced changes in host chemistry for plant productivity (Fig. 1).
Members of the microbiota inhibit pathogens through mechanisms such as chemical production and nutrient competition. They produce antimicrobial compounds that directly inhibit or kill pathogens, and secrete enzymes that degrade pathogen structures. For example, analysis of age-related microbiota in Populus tomentosa shows how mature trees develop more diverse microbial communities that are gradually enriched in Actinobacterial taxa (Xie et al., 2023). Corresponding isolates efficiently suppress tree diseases through direct antimicrobial-mediated pathogen suppression and therefore likely supplement host innate immunity to protect older trees in nature. Enrichment of host-protective taxa has also been observed in the context of crop rotation. Hong et al. (2023) revealed that crop rotation-induced suppression of banana wilt disease was associated with rotation-enriched microbial taxa that displayed strong antagonistic activity against Fusarium oxysporum pathogens and protected the host from disease. Additionally, microbiota members compete with pathogens for essential nutrients and space, effectively limiting access to pathogens and growth opportunities. For example, Zhou et al. (2023) tested the influence of carbon resource diversity on the microbial trophic network architecture and pathogen invasion success using tomato microcosm experiments. The authors concluded that expanding the carbon source diversity increased microbial evenness, widened the microbial functional landscape, and promoted disease suppression via efficient resource competition with pathogens (Zhou et al., 2023). Similarly, the critical function of host-derived sugars for bacterial niche competition in flowers and for the modulation of virulence factor expression in pathogens has recently been discussed by Lee et al. (2024). These authors observed that flower-derived arabinogalactan is an important sugar that drives pathogen colonization, and that competition for arabinogalactan between the pathogen and the sympatric commensal Pantoea agglomerans is likely key for P. agglomerans-mediated disease suppression. Furthermore, arabinogalactan consumption by P. agglomerans alters the production of major virulence determinants in the pathogen (Lee et al., 2024). These findings underscore the importance of microbe-mediated chemical and nutrient blocking for successful niche colonization and pathogen suppression (Fig. 1).
Over time, host-adapted pathogens have evolved strategies to manipulate microbiota to promote infection. Several reports present compelling evidence that filamentous plant pathogens can manipulate plant microbiota to their advantage by targeting beneficial microbial communities. Although pathogens are also known to alter microbiota assembly through chemical and nutrient competition, several reports published in this Virtual Issue indicate that they also use effector proteins and enzymes to do so. For instance, the Verticillium dahliae effector gene VdAve1L exhibits extensive allelic variation. Snelders et al. (2023) demonstrated that the full-length effector variant VdAve1L2 has antimicrobial activity, which the fungus uses to mediate tomato colonization by directly suppressing the antagonistic Actinobacteria in the host microbiota. Similarly, the parasitic protist Albugo candida releases proteins into the apoplast of the plant that selectively suppress beneficial phyllosphere-associated bacteria (gram-positive) (Gómez-Pérez et al., 2023; Rovenich & Thomma, 2023). These authors observed that intrinsically disordered regions of the protein and a high-positive net charge are often associated with the antibacterial activity of the proteins (Gómez-Pérez et al., 2023). Finally, another report revealed that the smut fungus Ustilago maydis exclusively expresses a gene encoding ribotoxin (Ribo1) at early infection stages (Ökmen et al., 2023). This secreted ribonuclease has a wide range of cytotoxic activities against bacteria and yeast cells and enables smut fungi to compete with host-associated bacteria. Taken together, these strategies enable pathogens to outcompete host-associated bacteria to enhance their colonization (Fig. 1).
In conclusion, we hope that this Virtual Issue nicely illustrates that chemical communication between organisms drives high-level biological organization and ecosystem functioning, and represents a key mechanism by which plants and microbes adapt to their often-stressful surrounding environments (Fig. 1). The remarkable diversity of molecules and mechanisms deployed by living organisms to respond to environmental cues is key for species survival and therefore contributes to landscape diversity.
期刊介绍:
New Phytologist is an international electronic journal published 24 times a year. It is owned by the New Phytologist Foundation, a non-profit-making charitable organization dedicated to promoting plant science. The journal publishes excellent, novel, rigorous, and timely research and scholarship in plant science and its applications. The articles cover topics in five sections: Physiology & Development, Environment, Interaction, Evolution, and Transformative Plant Biotechnology. These sections encompass intracellular processes, global environmental change, and encourage cross-disciplinary approaches. The journal recognizes the use of techniques from molecular and cell biology, functional genomics, modeling, and system-based approaches in plant science. Abstracting and Indexing Information for New Phytologist includes Academic Search, AgBiotech News & Information, Agroforestry Abstracts, Biochemistry & Biophysics Citation Index, Botanical Pesticides, CAB Abstracts®, Environment Index, Global Health, and Plant Breeding Abstracts, and others.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


