Timothy J. Fernandes, Reilly O'Connor, Kevin S. McCann, Brian J. Shuter, Bailey C. McMeans
{"title":"中食性太阳鱼的短暂食鱼行为:对池塘食物网的影响","authors":"Timothy J. Fernandes, Reilly O'Connor, Kevin S. McCann, Brian J. Shuter, Bailey C. McMeans","doi":"10.1002/ecy.4431","DOIUrl":null,"url":null,"abstract":"<p>Warm-water fishes are expanding northward rapidly across North America (Wu et al., <span>2023</span>), reshaping previously naïve north-temperate freshwater food webs (Vander Zanden et al., <span>1999</span>). In north-temperate lakes, large centrarchids (i.e., sunfishes) can drive declines in prey fish diversity and abundance and affect the foraging behavior of mesopredator fishes (e.g., introducing a landscape of “fear” that may restrict trophic breadth; Falkegård et al., <span>2023</span>). However, in pond systems where large predators are generally absent, we know relatively little about mesopredator ecology and food web interactions. Examining the ecological roles of mesopredators in ecosystems that lack large top predator species may reveal insights into their capacity to fill top predator niches (i.e., mesopredator release hypothesis). Yet, existing knowledge of mesopredator fish ecology is largely based on data collected in lakes, with limited work considering their potential role as top predators in pond ecosystems (Copp et al., <span>2017</span>; Fox & Keast, <span>1990</span>).</p><p>To better document the seasonal ecology of mesopredator fish populations across a range of pond habitats (Figure 1), we sampled four pond populations of pumpkinseed sunfish (<i>Lepomis gibbosus</i>) in Southern Ontario, Canada, biweekly from January to August (<i>N</i> = 763; Appendix S1: Table S1). Pumpkinseed sunfish are small, vibrantly colored centrarchids that are largely regarded as generalist mesopredators, consuming a range of dipterans, mollusks, gastropods, and pelagic zooplankton (Scott & Crossman, <span>1998</span>). This capacity for diverse resource polymorphisms combined with broad oxythermal preferences allows pumpkinseed to inhabit a range of aquatic habitats, from small ponds to the Great Lakes (Copp et al., <span>2017</span>; Scott & Crossman, <span>1998</span>), while also being among the most rapidly expanding fish species in Ontario (Wu et al., <span>2023</span>). To investigate seasonality in trophic interactions, pumpkinseed were sampled and dissected for diet analysis; it was during these dissections that we documented a unique seasonal interaction between pumpkinseed and small-bodied fishes. During late spring and summer months, pumpkinseed exhibited extensive seasonal piscivory in three of the four sampled ponds (Figure 1), consuming brook stickleback <i>Culaea inconstans</i>, fathead minnow <i>Pimephales pimelas</i>, northern redbelly dace <i>Chromosomus eos</i>, and young-of-the-year (YOY) pumpkinseed. According to the index of relative importance (Appendix S2: Equation S1; Hart et al., <span>2002</span>), fish became the most important diet item in Pond B and Pond C; though to a lesser extent, fish also emerged as an important prey category in Pond A (Appendix S1: Figure S1). The only pond where piscivory was not observed (Pond D) was also the only pond that contained a piscivorous predator (largemouth bass <i>Micropterus salmoides</i>).</p><p>Previous evidence, predominantly collected from lake populations, has suggested that fish consumption by pumpkinseed is exceedingly rare and limited to infrequent events involving only the largest individuals (>120–140 mm in total length; Collar et al., <span>2009</span>; Copp & Fox, <span>2007</span>; Rezsu & Specziár, <span>2006</span>). Though pumpkinseed have been assumed to consume leuciscid larvae (Godinho & Ferreira, <span>1998</span>), few observations of piscivory in this mesopredator have ever been recorded (Copp & Fox, <span>2007</span>; Rezsu & Specziár, <span>2006</span>). However, we demonstrate that piscivory in ponds can arise even in a small-bodied pumpkinseed population (Pond C) where individuals rarely reach lengths above 110 mm (Appendix S1: Table S2). For piscivorous pumpkinseed, estimates of predator–prey body mass ratios ranged from 1.1 to 2.6 (median = 1.5, <i>n</i> = 36; log<sub>10</sub> individual-link predator–prey mass ratio as in Gaeta et al., <span>2018</span>; see Appendix S2: Equation S2). Thus, regardless of expected gape limitation in pumpkinseed (Collar et al., <span>2009</span>), piscivorous individuals exhibited similar predator–prey body mass ratios to larger bodied freshwater piscivores (range of median values for <i>Poxomis</i> spp., <i>Micropterus</i> spp., <i>Sander</i> spp., and <i>Esox</i> spp. = 1.9–2.5; Gaeta et al., <span>2018</span>). When large piscivores were present (Pond D), pumpkinseed failed to consume fish prey, feeding exclusively on invertebrates. Though sample sizes from Pond D were relatively limited (<i>n</i> = 51; Appendix S1: Table S1), pumpkinseed are known to exhibit reduced trophic breadth in the presence of piscivores, including largemouth bass (Godinho et al., <span>1997</span>). Thus, release from top predators (i.e., mesopredator release) appears to encourage pumpkinseed, a generalist mesopredator, to fill the piscivorous top predator niche. We contend that the lack of consideration for fish in pond food webs and limited repeated seasonal sampling in these systems has prevented previous observations of piscivory within and across pumpkinseed populations.</p><p>Ponds commonly host abundant fish communities (Scheffer & van Geest, <span>2006</span>); however, these communities are largely comprised of small-bodied prey fishes (e.g., Leuciscidae) that are capable of achieving high densities in the absence of larger piscivores (e.g., smallmouth bass <i>M. dolomieu</i>). In lake habitats, small-bodied prey fishes exhibit contracting geographic distributions in response to predator range expansions (Wu et al., <span>2023</span>). Thus, pond habitats that are generally free of larger piscivores may be important yet poorly documented source populations for small forage fish species, like leuciscids. These same habitats may offer unique opportunities for mesopredator release, as observed here in pumpkinseed that appeared to expand their niche to exploit higher trophic level prey in the absence of coexisting piscivores. Indeed, previous work has suggested that pond-resident pumpkinseed exhibit greater diet diversity than populations found in other aquatic habitats (Haubrock et al., <span>2021</span>). Further work should consider using pond ecosystems to more explicitly test the mesopredator release hypothesis, leveraging ponds with divergent predator communities and investigating outcomes across multiple trophic levels (e.g., primary productivity, invertebrate abundance, diversity, foraging ecology, food chain length). Historically, the impacts of range expansions on aquatic food webs have been focused on top predator species (Alofs & Jackson, <span>2015</span>; Vander Zanden et al., <span>1999</span>). However, evidence here underscores the potential yet underappreciated consequences that the expansion of mesopredator species, like pumpkinseed, may have for aquatic ecosystems. Recent evidence also suggests that mesopredators may be the most rapidly expanding trophic guild of freshwater fishes, further substantiating a need for focused future work on mesopredators in pond food webs (Wu et al., <span>2023</span>).</p><p>In invaded north-temperate lakes, smallmouth bass reduce forage fish abundance and alter coexisting predator behavior (Vander Zanden et al., <span>1999</span>). Similarly, introduced pumpkinseed populations in Europe drove contractions in the overall niche space and reduced body condition and growth in native small-bodied fishes (Copp et al., <span>2017</span>). Though we lack information on the origin and colonization history of each pond sampled here, pumpkinseed likely exert similar ecological pressures on native forage fishes in North American ponds. In this way, ponds may represent systems that are particularly sensitive to invasion, analogous to small aquatic islands (Elton, <span>2020</span>). Similar to the consequences of novel predators colonizing isolated islands (e.g., brown tree snakes <i>Boiga irregularis</i> depredating native avifauna in Guam; Wiles et al., <span>2003</span>), the expansion of sunfish and other predatory fishes through pond networks may have similarly drastic consequences for aquatic food web stability and diversity in these forgotten yet invaluable ecosystems. Though pumpkinseed are capable of reducing the abundance and diversity of macroinvertebrate communities in invaded systems (Van Kleef et al., <span>2008</span>), how predation of smaller invertivorous fishes may cascade to reshape pond food webs is currently unclear. The seasonal patterning of fish consumption by pumpkinseed here also merits further discussion.</p><p>In two of the ponds where piscivory was observed (Pond A, Pond B), pumpkinseed co-occurred with both leuciscid and stickleback species (Appendix S1: Table S2). Dace and fathead minnows were consumed most heavily when they were actively spawning and nest-guarding (late May to late July; T. Fernandes, personal observation). These prey fish species are particularly vulnerable to predation when spawning and nest guarding (Jones & Paszkowski, <span>1997</span>), potentially facilitating predation by pumpkinseed during these periods. In Pond C, where no other fish species were present, mature though small-bodied pumpkinseed became cannibalistic and consumed YOY fish nearly exclusively during summer (Figure 1E). In small, less productive habitats, cannibalism may represent an important mechanism for density-dependent control (Claessen et al., <span>2004</span>), while contributing a valuable pulse of energy when little else may be available.</p><p>Here, we document seasonal piscivory across populations of a widespread and rapidly expanding warm-water sunfish, the pumpkinseed. These observations raise important questions about ecological theory and aquatic food webs and highlight the value of seasonal sampling for uncovering novel ephemeral interactions. Furthermore, they underscore the need for continued consideration of ponds as important systems in which to test ecological hypotheses and quantify the potential impacts of rapidly expanding mesopredators. Future work would benefit from more explicitly testing the mesopredator release hypothesis by considering a wider array of pond habitats that contain varied fish predator communities. By considering ponds that range is size, this work could also employ novel tests of the ecosystem size and productive-space hypotheses for predicting food web properties (e.g., food chain length, predator–prey mass rations). In addition, further work is needed to quantify the consequences of both meso- and top predator expansion into pond food webs, as ponds may buffer the risk of regional declines in small-bodied fishes and offer practical models for predicting consequences in larger systems. In a rapidly changing world, understanding novel interactions that have the potential to rewire food webs, whether found in the smallest of ponds to the largest of lakes, will be vital for managing resilience and the persistence of biodiversity across connected landscapes.</p><p>Timothy J. Fernandes, Brian J. Shuter, and Bailey C. McMeans conceived the study design. Timothy J. Fernandes conducted fieldwork, dissections, and dataset curation, assisted with data analysis, and co-wrote the manuscript. Reilly O'Connor and Kevin S. McCann curated and analyzed the data and co-wrote the manuscript. All authors contributed to the writing of the manuscript and the ideas put forth.</p><p>The authors declare no conflicts of interest.</p>","PeriodicalId":11484,"journal":{"name":"Ecology","volume":"105 11","pages":""},"PeriodicalIF":4.4000,"publicationDate":"2024-10-01","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1002/ecy.4431","citationCount":"0","resultStr":"{\"title\":\"Ephemeral piscivory in a mesopredator sunfish: Implications for pond food webs\",\"authors\":\"Timothy J. Fernandes, Reilly O'Connor, Kevin S. McCann, Brian J. Shuter, Bailey C. McMeans\",\"doi\":\"10.1002/ecy.4431\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p>Warm-water fishes are expanding northward rapidly across North America (Wu et al., <span>2023</span>), reshaping previously naïve north-temperate freshwater food webs (Vander Zanden et al., <span>1999</span>). In north-temperate lakes, large centrarchids (i.e., sunfishes) can drive declines in prey fish diversity and abundance and affect the foraging behavior of mesopredator fishes (e.g., introducing a landscape of “fear” that may restrict trophic breadth; Falkegård et al., <span>2023</span>). However, in pond systems where large predators are generally absent, we know relatively little about mesopredator ecology and food web interactions. Examining the ecological roles of mesopredators in ecosystems that lack large top predator species may reveal insights into their capacity to fill top predator niches (i.e., mesopredator release hypothesis). Yet, existing knowledge of mesopredator fish ecology is largely based on data collected in lakes, with limited work considering their potential role as top predators in pond ecosystems (Copp et al., <span>2017</span>; Fox & Keast, <span>1990</span>).</p><p>To better document the seasonal ecology of mesopredator fish populations across a range of pond habitats (Figure 1), we sampled four pond populations of pumpkinseed sunfish (<i>Lepomis gibbosus</i>) in Southern Ontario, Canada, biweekly from January to August (<i>N</i> = 763; Appendix S1: Table S1). Pumpkinseed sunfish are small, vibrantly colored centrarchids that are largely regarded as generalist mesopredators, consuming a range of dipterans, mollusks, gastropods, and pelagic zooplankton (Scott & Crossman, <span>1998</span>). This capacity for diverse resource polymorphisms combined with broad oxythermal preferences allows pumpkinseed to inhabit a range of aquatic habitats, from small ponds to the Great Lakes (Copp et al., <span>2017</span>; Scott & Crossman, <span>1998</span>), while also being among the most rapidly expanding fish species in Ontario (Wu et al., <span>2023</span>). To investigate seasonality in trophic interactions, pumpkinseed were sampled and dissected for diet analysis; it was during these dissections that we documented a unique seasonal interaction between pumpkinseed and small-bodied fishes. During late spring and summer months, pumpkinseed exhibited extensive seasonal piscivory in three of the four sampled ponds (Figure 1), consuming brook stickleback <i>Culaea inconstans</i>, fathead minnow <i>Pimephales pimelas</i>, northern redbelly dace <i>Chromosomus eos</i>, and young-of-the-year (YOY) pumpkinseed. According to the index of relative importance (Appendix S2: Equation S1; Hart et al., <span>2002</span>), fish became the most important diet item in Pond B and Pond C; though to a lesser extent, fish also emerged as an important prey category in Pond A (Appendix S1: Figure S1). The only pond where piscivory was not observed (Pond D) was also the only pond that contained a piscivorous predator (largemouth bass <i>Micropterus salmoides</i>).</p><p>Previous evidence, predominantly collected from lake populations, has suggested that fish consumption by pumpkinseed is exceedingly rare and limited to infrequent events involving only the largest individuals (>120–140 mm in total length; Collar et al., <span>2009</span>; Copp & Fox, <span>2007</span>; Rezsu & Specziár, <span>2006</span>). Though pumpkinseed have been assumed to consume leuciscid larvae (Godinho & Ferreira, <span>1998</span>), few observations of piscivory in this mesopredator have ever been recorded (Copp & Fox, <span>2007</span>; Rezsu & Specziár, <span>2006</span>). However, we demonstrate that piscivory in ponds can arise even in a small-bodied pumpkinseed population (Pond C) where individuals rarely reach lengths above 110 mm (Appendix S1: Table S2). For piscivorous pumpkinseed, estimates of predator–prey body mass ratios ranged from 1.1 to 2.6 (median = 1.5, <i>n</i> = 36; log<sub>10</sub> individual-link predator–prey mass ratio as in Gaeta et al., <span>2018</span>; see Appendix S2: Equation S2). Thus, regardless of expected gape limitation in pumpkinseed (Collar et al., <span>2009</span>), piscivorous individuals exhibited similar predator–prey body mass ratios to larger bodied freshwater piscivores (range of median values for <i>Poxomis</i> spp., <i>Micropterus</i> spp., <i>Sander</i> spp., and <i>Esox</i> spp. = 1.9–2.5; Gaeta et al., <span>2018</span>). When large piscivores were present (Pond D), pumpkinseed failed to consume fish prey, feeding exclusively on invertebrates. Though sample sizes from Pond D were relatively limited (<i>n</i> = 51; Appendix S1: Table S1), pumpkinseed are known to exhibit reduced trophic breadth in the presence of piscivores, including largemouth bass (Godinho et al., <span>1997</span>). Thus, release from top predators (i.e., mesopredator release) appears to encourage pumpkinseed, a generalist mesopredator, to fill the piscivorous top predator niche. We contend that the lack of consideration for fish in pond food webs and limited repeated seasonal sampling in these systems has prevented previous observations of piscivory within and across pumpkinseed populations.</p><p>Ponds commonly host abundant fish communities (Scheffer & van Geest, <span>2006</span>); however, these communities are largely comprised of small-bodied prey fishes (e.g., Leuciscidae) that are capable of achieving high densities in the absence of larger piscivores (e.g., smallmouth bass <i>M. dolomieu</i>). In lake habitats, small-bodied prey fishes exhibit contracting geographic distributions in response to predator range expansions (Wu et al., <span>2023</span>). Thus, pond habitats that are generally free of larger piscivores may be important yet poorly documented source populations for small forage fish species, like leuciscids. These same habitats may offer unique opportunities for mesopredator release, as observed here in pumpkinseed that appeared to expand their niche to exploit higher trophic level prey in the absence of coexisting piscivores. Indeed, previous work has suggested that pond-resident pumpkinseed exhibit greater diet diversity than populations found in other aquatic habitats (Haubrock et al., <span>2021</span>). Further work should consider using pond ecosystems to more explicitly test the mesopredator release hypothesis, leveraging ponds with divergent predator communities and investigating outcomes across multiple trophic levels (e.g., primary productivity, invertebrate abundance, diversity, foraging ecology, food chain length). Historically, the impacts of range expansions on aquatic food webs have been focused on top predator species (Alofs & Jackson, <span>2015</span>; Vander Zanden et al., <span>1999</span>). However, evidence here underscores the potential yet underappreciated consequences that the expansion of mesopredator species, like pumpkinseed, may have for aquatic ecosystems. Recent evidence also suggests that mesopredators may be the most rapidly expanding trophic guild of freshwater fishes, further substantiating a need for focused future work on mesopredators in pond food webs (Wu et al., <span>2023</span>).</p><p>In invaded north-temperate lakes, smallmouth bass reduce forage fish abundance and alter coexisting predator behavior (Vander Zanden et al., <span>1999</span>). Similarly, introduced pumpkinseed populations in Europe drove contractions in the overall niche space and reduced body condition and growth in native small-bodied fishes (Copp et al., <span>2017</span>). Though we lack information on the origin and colonization history of each pond sampled here, pumpkinseed likely exert similar ecological pressures on native forage fishes in North American ponds. In this way, ponds may represent systems that are particularly sensitive to invasion, analogous to small aquatic islands (Elton, <span>2020</span>). Similar to the consequences of novel predators colonizing isolated islands (e.g., brown tree snakes <i>Boiga irregularis</i> depredating native avifauna in Guam; Wiles et al., <span>2003</span>), the expansion of sunfish and other predatory fishes through pond networks may have similarly drastic consequences for aquatic food web stability and diversity in these forgotten yet invaluable ecosystems. Though pumpkinseed are capable of reducing the abundance and diversity of macroinvertebrate communities in invaded systems (Van Kleef et al., <span>2008</span>), how predation of smaller invertivorous fishes may cascade to reshape pond food webs is currently unclear. The seasonal patterning of fish consumption by pumpkinseed here also merits further discussion.</p><p>In two of the ponds where piscivory was observed (Pond A, Pond B), pumpkinseed co-occurred with both leuciscid and stickleback species (Appendix S1: Table S2). Dace and fathead minnows were consumed most heavily when they were actively spawning and nest-guarding (late May to late July; T. Fernandes, personal observation). These prey fish species are particularly vulnerable to predation when spawning and nest guarding (Jones & Paszkowski, <span>1997</span>), potentially facilitating predation by pumpkinseed during these periods. In Pond C, where no other fish species were present, mature though small-bodied pumpkinseed became cannibalistic and consumed YOY fish nearly exclusively during summer (Figure 1E). In small, less productive habitats, cannibalism may represent an important mechanism for density-dependent control (Claessen et al., <span>2004</span>), while contributing a valuable pulse of energy when little else may be available.</p><p>Here, we document seasonal piscivory across populations of a widespread and rapidly expanding warm-water sunfish, the pumpkinseed. These observations raise important questions about ecological theory and aquatic food webs and highlight the value of seasonal sampling for uncovering novel ephemeral interactions. Furthermore, they underscore the need for continued consideration of ponds as important systems in which to test ecological hypotheses and quantify the potential impacts of rapidly expanding mesopredators. Future work would benefit from more explicitly testing the mesopredator release hypothesis by considering a wider array of pond habitats that contain varied fish predator communities. By considering ponds that range is size, this work could also employ novel tests of the ecosystem size and productive-space hypotheses for predicting food web properties (e.g., food chain length, predator–prey mass rations). In addition, further work is needed to quantify the consequences of both meso- and top predator expansion into pond food webs, as ponds may buffer the risk of regional declines in small-bodied fishes and offer practical models for predicting consequences in larger systems. In a rapidly changing world, understanding novel interactions that have the potential to rewire food webs, whether found in the smallest of ponds to the largest of lakes, will be vital for managing resilience and the persistence of biodiversity across connected landscapes.</p><p>Timothy J. Fernandes, Brian J. Shuter, and Bailey C. McMeans conceived the study design. Timothy J. Fernandes conducted fieldwork, dissections, and dataset curation, assisted with data analysis, and co-wrote the manuscript. Reilly O'Connor and Kevin S. McCann curated and analyzed the data and co-wrote the manuscript. All authors contributed to the writing of the manuscript and the ideas put forth.</p><p>The authors declare no conflicts of interest.</p>\",\"PeriodicalId\":11484,\"journal\":{\"name\":\"Ecology\",\"volume\":\"105 11\",\"pages\":\"\"},\"PeriodicalIF\":4.4000,\"publicationDate\":\"2024-10-01\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://onlinelibrary.wiley.com/doi/epdf/10.1002/ecy.4431\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"Ecology\",\"FirstCategoryId\":\"93\",\"ListUrlMain\":\"https://onlinelibrary.wiley.com/doi/10.1002/ecy.4431\",\"RegionNum\":2,\"RegionCategory\":\"环境科学与生态学\",\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"Q1\",\"JCRName\":\"ECOLOGY\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"Ecology","FirstCategoryId":"93","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1002/ecy.4431","RegionNum":2,"RegionCategory":"环境科学与生态学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"ECOLOGY","Score":null,"Total":0}
引用次数: 0
摘要
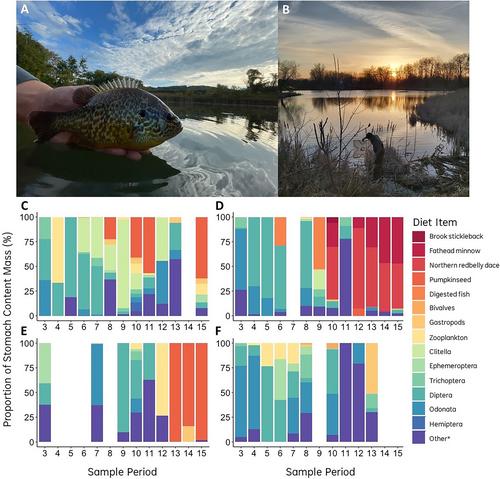
池塘通常拥有丰富的鱼类群落(Scheffer & van Geest, 2006);然而,这些群落主要由小型猎食鱼类(如鯈鱼)组成,它们能够在没有大型食鱼动物(如小口鲈)的情况下达到较高的密度。在湖泊生境中,小体型猎物鱼类的地理分布会随着捕食者范围的扩大而收缩(Wu 等,2023 年)。因此,一般没有大型食鱼动物的池塘栖息地可能是小型饵料鱼类(如白鲦鱼)的重要来源种群,但却鲜有记录。这些栖息地也可能为中型食肉动物的释放提供了独特的机会,就像这里观察到的南瓜籽,在没有共存的食鱼动物的情况下,它们似乎扩大了自己的生态位,以捕食更高营养级的猎物。事实上,之前的研究表明,与其他水生栖息地的种群相比,池塘栖息的南瓜籽表现出更大的食物多样性(Haubrock等人,2021年)。进一步的工作应考虑利用池塘生态系统来更明确地检验中间捕食者释放假说,利用具有不同捕食者群落的池塘,调查多个营养级(如初级生产力、无脊椎动物丰度、多样性、觅食生态学、食物链长度)的结果。从历史上看,范围扩大对水生食物网的影响主要集中在顶级捕食者物种上(Alofs & Jackson, 2015; Vander Zanden et al.)然而,这里的证据强调了南瓜籽等中层食肉动物的扩张可能对水生生态系统造成的潜在但未被充分认识的后果。最近的证据还表明,中食性鱼类可能是淡水鱼类中扩张最快的营养联盟,这进一步证实了今后需要重点研究池塘食物网中的中食性鱼类(Wu 等人,2023 年)。在被入侵的北温带湖泊中,小口鲈鱼会减少饲料鱼的数量,并改变共存的捕食者行为(Vander Zanden 等人,1999 年)。同样,欧洲引入的南瓜籽种群导致整体生态位空间收缩,并降低了本地小体型鱼类的身体状况和生长速度(Copp 等人,2017 年)。虽然我们缺乏有关每个取样池塘的起源和殖民历史的信息,但南瓜籽可能对北美池塘中的本地饲料鱼类施加了类似的生态压力。因此,池塘可能是对入侵特别敏感的系统,类似于小型水生岛屿(Elton,2020 年)。与新捕食者在孤立岛屿上殖民的后果类似(例如,关岛的棕色树蛇(Boiga irregularis)捕食本地鸟类;Wiles 等人,2003 年),太阳鱼和其他捕食性鱼类通过池塘网络的扩张可能会对这些被遗忘但却宝贵的生态系统中的水生食物网的稳定性和多样性产生类似的严重后果。虽然南瓜籽能够减少受入侵系统中大型无脊椎动物群落的数量和多样性(Van Kleef 等人,2008 年),但目前还不清楚小型无脊椎鱼类的捕食会如何连锁重塑池塘食物网。在观察到鱼类捕食的两个池塘(池塘 A、池塘 B)中,南瓜籽与白鲦鱼和竹刀鱼共生(附录 S1:表 S2)。鲂鱼和胖头鲦鱼在产卵和护巢期间(5 月下旬至 7 月下旬;T. Fernandes,个人观察)被大量捕食。这些猎物鱼类在产卵和护巢时特别容易被捕食(Jones & Paszkowski, 1997),可能会在这些时期促进南瓜籽的捕食。在没有其他鱼类的池塘 C 中,成熟但体型较小的南瓜鱼开始食人,几乎只在夏季捕食幼鱼(图 1E)。在小型、生产力较低的栖息地,食人可能是一种重要的密度控制机制(Claessen 等,2004 年),同时在几乎没有其他鱼类的情况下提供宝贵的能量脉冲。这些观察结果提出了有关生态理论和水生食物网的重要问题,并强调了季节性取样在发现新的短暂相互作用方面的价值。此外,它们还强调了继续将池塘作为重要系统的必要性,以检验生态假说并量化快速扩张的中型食肉动物的潜在影响。通过考虑包含不同鱼类捕食者群落的更广泛池塘栖息地,更明确地测试中型捕食者释放假说将有利于未来的工作。
Ephemeral piscivory in a mesopredator sunfish: Implications for pond food webs
Warm-water fishes are expanding northward rapidly across North America (Wu et al., 2023), reshaping previously naïve north-temperate freshwater food webs (Vander Zanden et al., 1999). In north-temperate lakes, large centrarchids (i.e., sunfishes) can drive declines in prey fish diversity and abundance and affect the foraging behavior of mesopredator fishes (e.g., introducing a landscape of “fear” that may restrict trophic breadth; Falkegård et al., 2023). However, in pond systems where large predators are generally absent, we know relatively little about mesopredator ecology and food web interactions. Examining the ecological roles of mesopredators in ecosystems that lack large top predator species may reveal insights into their capacity to fill top predator niches (i.e., mesopredator release hypothesis). Yet, existing knowledge of mesopredator fish ecology is largely based on data collected in lakes, with limited work considering their potential role as top predators in pond ecosystems (Copp et al., 2017; Fox & Keast, 1990).
To better document the seasonal ecology of mesopredator fish populations across a range of pond habitats (Figure 1), we sampled four pond populations of pumpkinseed sunfish (Lepomis gibbosus) in Southern Ontario, Canada, biweekly from January to August (N = 763; Appendix S1: Table S1). Pumpkinseed sunfish are small, vibrantly colored centrarchids that are largely regarded as generalist mesopredators, consuming a range of dipterans, mollusks, gastropods, and pelagic zooplankton (Scott & Crossman, 1998). This capacity for diverse resource polymorphisms combined with broad oxythermal preferences allows pumpkinseed to inhabit a range of aquatic habitats, from small ponds to the Great Lakes (Copp et al., 2017; Scott & Crossman, 1998), while also being among the most rapidly expanding fish species in Ontario (Wu et al., 2023). To investigate seasonality in trophic interactions, pumpkinseed were sampled and dissected for diet analysis; it was during these dissections that we documented a unique seasonal interaction between pumpkinseed and small-bodied fishes. During late spring and summer months, pumpkinseed exhibited extensive seasonal piscivory in three of the four sampled ponds (Figure 1), consuming brook stickleback Culaea inconstans, fathead minnow Pimephales pimelas, northern redbelly dace Chromosomus eos, and young-of-the-year (YOY) pumpkinseed. According to the index of relative importance (Appendix S2: Equation S1; Hart et al., 2002), fish became the most important diet item in Pond B and Pond C; though to a lesser extent, fish also emerged as an important prey category in Pond A (Appendix S1: Figure S1). The only pond where piscivory was not observed (Pond D) was also the only pond that contained a piscivorous predator (largemouth bass Micropterus salmoides).
Previous evidence, predominantly collected from lake populations, has suggested that fish consumption by pumpkinseed is exceedingly rare and limited to infrequent events involving only the largest individuals (>120–140 mm in total length; Collar et al., 2009; Copp & Fox, 2007; Rezsu & Specziár, 2006). Though pumpkinseed have been assumed to consume leuciscid larvae (Godinho & Ferreira, 1998), few observations of piscivory in this mesopredator have ever been recorded (Copp & Fox, 2007; Rezsu & Specziár, 2006). However, we demonstrate that piscivory in ponds can arise even in a small-bodied pumpkinseed population (Pond C) where individuals rarely reach lengths above 110 mm (Appendix S1: Table S2). For piscivorous pumpkinseed, estimates of predator–prey body mass ratios ranged from 1.1 to 2.6 (median = 1.5, n = 36; log10 individual-link predator–prey mass ratio as in Gaeta et al., 2018; see Appendix S2: Equation S2). Thus, regardless of expected gape limitation in pumpkinseed (Collar et al., 2009), piscivorous individuals exhibited similar predator–prey body mass ratios to larger bodied freshwater piscivores (range of median values for Poxomis spp., Micropterus spp., Sander spp., and Esox spp. = 1.9–2.5; Gaeta et al., 2018). When large piscivores were present (Pond D), pumpkinseed failed to consume fish prey, feeding exclusively on invertebrates. Though sample sizes from Pond D were relatively limited (n = 51; Appendix S1: Table S1), pumpkinseed are known to exhibit reduced trophic breadth in the presence of piscivores, including largemouth bass (Godinho et al., 1997). Thus, release from top predators (i.e., mesopredator release) appears to encourage pumpkinseed, a generalist mesopredator, to fill the piscivorous top predator niche. We contend that the lack of consideration for fish in pond food webs and limited repeated seasonal sampling in these systems has prevented previous observations of piscivory within and across pumpkinseed populations.
Ponds commonly host abundant fish communities (Scheffer & van Geest, 2006); however, these communities are largely comprised of small-bodied prey fishes (e.g., Leuciscidae) that are capable of achieving high densities in the absence of larger piscivores (e.g., smallmouth bass M. dolomieu). In lake habitats, small-bodied prey fishes exhibit contracting geographic distributions in response to predator range expansions (Wu et al., 2023). Thus, pond habitats that are generally free of larger piscivores may be important yet poorly documented source populations for small forage fish species, like leuciscids. These same habitats may offer unique opportunities for mesopredator release, as observed here in pumpkinseed that appeared to expand their niche to exploit higher trophic level prey in the absence of coexisting piscivores. Indeed, previous work has suggested that pond-resident pumpkinseed exhibit greater diet diversity than populations found in other aquatic habitats (Haubrock et al., 2021). Further work should consider using pond ecosystems to more explicitly test the mesopredator release hypothesis, leveraging ponds with divergent predator communities and investigating outcomes across multiple trophic levels (e.g., primary productivity, invertebrate abundance, diversity, foraging ecology, food chain length). Historically, the impacts of range expansions on aquatic food webs have been focused on top predator species (Alofs & Jackson, 2015; Vander Zanden et al., 1999). However, evidence here underscores the potential yet underappreciated consequences that the expansion of mesopredator species, like pumpkinseed, may have for aquatic ecosystems. Recent evidence also suggests that mesopredators may be the most rapidly expanding trophic guild of freshwater fishes, further substantiating a need for focused future work on mesopredators in pond food webs (Wu et al., 2023).
In invaded north-temperate lakes, smallmouth bass reduce forage fish abundance and alter coexisting predator behavior (Vander Zanden et al., 1999). Similarly, introduced pumpkinseed populations in Europe drove contractions in the overall niche space and reduced body condition and growth in native small-bodied fishes (Copp et al., 2017). Though we lack information on the origin and colonization history of each pond sampled here, pumpkinseed likely exert similar ecological pressures on native forage fishes in North American ponds. In this way, ponds may represent systems that are particularly sensitive to invasion, analogous to small aquatic islands (Elton, 2020). Similar to the consequences of novel predators colonizing isolated islands (e.g., brown tree snakes Boiga irregularis depredating native avifauna in Guam; Wiles et al., 2003), the expansion of sunfish and other predatory fishes through pond networks may have similarly drastic consequences for aquatic food web stability and diversity in these forgotten yet invaluable ecosystems. Though pumpkinseed are capable of reducing the abundance and diversity of macroinvertebrate communities in invaded systems (Van Kleef et al., 2008), how predation of smaller invertivorous fishes may cascade to reshape pond food webs is currently unclear. The seasonal patterning of fish consumption by pumpkinseed here also merits further discussion.
In two of the ponds where piscivory was observed (Pond A, Pond B), pumpkinseed co-occurred with both leuciscid and stickleback species (Appendix S1: Table S2). Dace and fathead minnows were consumed most heavily when they were actively spawning and nest-guarding (late May to late July; T. Fernandes, personal observation). These prey fish species are particularly vulnerable to predation when spawning and nest guarding (Jones & Paszkowski, 1997), potentially facilitating predation by pumpkinseed during these periods. In Pond C, where no other fish species were present, mature though small-bodied pumpkinseed became cannibalistic and consumed YOY fish nearly exclusively during summer (Figure 1E). In small, less productive habitats, cannibalism may represent an important mechanism for density-dependent control (Claessen et al., 2004), while contributing a valuable pulse of energy when little else may be available.
Here, we document seasonal piscivory across populations of a widespread and rapidly expanding warm-water sunfish, the pumpkinseed. These observations raise important questions about ecological theory and aquatic food webs and highlight the value of seasonal sampling for uncovering novel ephemeral interactions. Furthermore, they underscore the need for continued consideration of ponds as important systems in which to test ecological hypotheses and quantify the potential impacts of rapidly expanding mesopredators. Future work would benefit from more explicitly testing the mesopredator release hypothesis by considering a wider array of pond habitats that contain varied fish predator communities. By considering ponds that range is size, this work could also employ novel tests of the ecosystem size and productive-space hypotheses for predicting food web properties (e.g., food chain length, predator–prey mass rations). In addition, further work is needed to quantify the consequences of both meso- and top predator expansion into pond food webs, as ponds may buffer the risk of regional declines in small-bodied fishes and offer practical models for predicting consequences in larger systems. In a rapidly changing world, understanding novel interactions that have the potential to rewire food webs, whether found in the smallest of ponds to the largest of lakes, will be vital for managing resilience and the persistence of biodiversity across connected landscapes.
Timothy J. Fernandes, Brian J. Shuter, and Bailey C. McMeans conceived the study design. Timothy J. Fernandes conducted fieldwork, dissections, and dataset curation, assisted with data analysis, and co-wrote the manuscript. Reilly O'Connor and Kevin S. McCann curated and analyzed the data and co-wrote the manuscript. All authors contributed to the writing of the manuscript and the ideas put forth.
期刊介绍:
Ecology publishes articles that report on the basic elements of ecological research. Emphasis is placed on concise, clear articles documenting important ecological phenomena. The journal publishes a broad array of research that includes a rapidly expanding envelope of subject matter, techniques, approaches, and concepts: paleoecology through present-day phenomena; evolutionary, population, physiological, community, and ecosystem ecology, as well as biogeochemistry; inclusive of descriptive, comparative, experimental, mathematical, statistical, and interdisciplinary approaches.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


