Andrew Barnas, Brad Anholt, A. Cole Burton, Kathleen Carroll, Steeve D. Côté, Marco Festa-Bianchet, John Fryxell, Martin-Hugues St-Laurent, Jason T. Fisher
{"title":"不应忽视生境改变对入侵白尾鹿密度的影响","authors":"Andrew Barnas, Brad Anholt, A. Cole Burton, Kathleen Carroll, Steeve D. Côté, Marco Festa-Bianchet, John Fryxell, Martin-Hugues St-Laurent, Jason T. Fisher","doi":"10.1111/gcb.17498","DOIUrl":null,"url":null,"abstract":"<p>White-tailed deer (<i>Odocoileus virginianus</i>) range expansion into boreal forests facilitates wolf (<i>Canis lupus</i>) population growth in many parts of Canada and is associated with caribou (<i>Rangifer tarandus caribou</i>) declines (Latham et al., <span>2011</span>). Several works across Canada have demonstrated anthropogenic landscape change subsidizes forage available to white-tailed deer (Darlington et al., <span>2022</span>; Fisher et al., <span>2020</span>). However, recently Dickie et al. (<span>2024</span>) suggested instead climate is the primary driver of spatial variation in white-tailed deer density in Canada's boreal forest. These findings garnered significant media attention (CBC, <span>2024</span>; NPR, <span>2024</span>) with direct implications for conservation actions, specifically habitat protection and restoration efforts. We generally agree with the authors' conclusion that spatial variation in winter severity impacts deer population density, but we contend their conclusion on negligible impact of habitat alteration was compromised by their method of transforming explanatory variables and insufficient data for their model.</p><p>The resulting coefficients from regression analyses with min-max scaled variables are interpreted as the change in the response variable, with an increase in the explanatory variable from the lowest to highest observed value.</p><p>Here, variables are transformed to quantify the difference of each observation from the mean in terms of standard deviations, whereby the data distribution is scaled to a mean of zero and standard deviation of one. Regression coefficients from z-standardized data represent the estimated change in the response variable per one unit increase in explanatory variables, where one unit represents one standard deviation in the z-standardized data.</p><p>Both transformations improve direct comparison between regression coefficients for variables measured on different scales, but differences in the algebraic transformation have considerable impact on conclusions. To demonstrate that Dickie et al. (<span>2024</span>)'s conclusions are sensitive to transformations, we z-standardized their original data and re-ran their global model examining effects of several variables on white-tailed deer density. Using min-max scaling, the authors originally reported negative statistically significant effects of Climate Dimension 1 (<i>β</i> = −6.794 ± 2.523, <i>p</i> < .007) and negative but insignificant effect of % Habitat Alteration (<i>β</i> = −0.328 ± 2.060, P = 0.873) (their Table 2). However, when using z-standardized data, we found a different conclusion of approximately equal magnitude but opposing effects of Climate Dimension 1 (<i>β</i> = −0.907 ± 0.286, <i>p</i> < .002) and % Habitat Alteration (<i>β</i> = 0.926 ± 0.307, <i>p</i> < .003, Figure 1).</p><p>Importantly, Dickie et al. (<span>2024</span>) describe a large dataset: “<i>…300 remote cameras across 12 replicated 50km</i><sup><i>2</i></sup> <i>landscapes over 5 years</i>.” and, “<i>Camera traps operated between 53,506 and 96,096 trap days from 2017 to 2021</i>”. However, the authors collapsed their response variable into a single mean density estimate for each camera cluster per year. The global model containing seven fixed effects (intercept, three “direct” effects, and three interactions), and two random effects, relies on only 53 data points. With a generous guideline of a minimum of 10 observations per treatment (Bolker et al., <span>2009</span>), their model has insufficient data to robustly estimate parameters of interest. This is likely the cause of large standard errors reported in their Table 2. The authors do admit estimates are imprecise, but do little to temper interpretations on climate vs. habitat alteration. We therefore constructed reduced models, which corroborate approximately equal but opposite effects of climate and habitat alteration on predicted deer density (Supplemental Materials S1).</p><p>Our reanalysis suggests both climate and habitat alteration drive density of white-tailed deer, but more extensive data across their Canadian range is required to estimate this properly. Parsing out the relative and interacting roles of these variables will be notoriously difficult at the landscape scale. Our reanalysis illustrates inherent risk of drawing potentially erroneous conclusions following different analytical decisions that produce contrasting results (Gould et al. <span>2023</span>). This is particularly problematic when conclusions are presented in social debates with direct impacts on conservation decisions by government authorities, as in woodland caribou conservation (CBC, <span>2024</span>). Given effective conservation actions depends on high-quality evidence, claims of which variables do and do not affect density of white-tailed deer in the boreal forest based on specific analytical decisions must be scrutinized.</p><p><b>Andrew Barnas:</b> Conceptualization; formal analysis; writing – original draft; writing – review and editing. <b>Brad Anholt:</b> Conceptualization; writing – original draft; writing – review and editing. <b>A. Cole Burton:</b> Conceptualization; writing – original draft; writing – review and editing. <b>Kathleen Carroll:</b> Conceptualization; writing – original draft; writing – review and editing. <b>Steeve D. Côté:</b> Conceptualization; writing – original draft; writing – review and editing. <b>Marco Festa-Bianchet:</b> Conceptualization; writing – original draft; writing – review and editing. <b>John Fryxell:</b> Conceptualization; writing – original draft; writing – review and editing. <b>Martin-Hugues St-Laurent:</b> Conceptualization; writing – original draft; writing – review and editing. <b>Jason T. Fisher:</b> Conceptualization; formal analysis; writing – original draft; writing – review and editing.</p><p>We have no conflict of interest to declare.</p>","PeriodicalId":175,"journal":{"name":"Global Change Biology","volume":"30 9","pages":""},"PeriodicalIF":10.8000,"publicationDate":"2024-09-13","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1111/gcb.17498","citationCount":"0","resultStr":"{\"title\":\"The influence of habitat alteration on density of invading white-tailed deer should not be discounted\",\"authors\":\"Andrew Barnas, Brad Anholt, A. Cole Burton, Kathleen Carroll, Steeve D. Côté, Marco Festa-Bianchet, John Fryxell, Martin-Hugues St-Laurent, Jason T. Fisher\",\"doi\":\"10.1111/gcb.17498\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p>White-tailed deer (<i>Odocoileus virginianus</i>) range expansion into boreal forests facilitates wolf (<i>Canis lupus</i>) population growth in many parts of Canada and is associated with caribou (<i>Rangifer tarandus caribou</i>) declines (Latham et al., <span>2011</span>). Several works across Canada have demonstrated anthropogenic landscape change subsidizes forage available to white-tailed deer (Darlington et al., <span>2022</span>; Fisher et al., <span>2020</span>). However, recently Dickie et al. (<span>2024</span>) suggested instead climate is the primary driver of spatial variation in white-tailed deer density in Canada's boreal forest. These findings garnered significant media attention (CBC, <span>2024</span>; NPR, <span>2024</span>) with direct implications for conservation actions, specifically habitat protection and restoration efforts. We generally agree with the authors' conclusion that spatial variation in winter severity impacts deer population density, but we contend their conclusion on negligible impact of habitat alteration was compromised by their method of transforming explanatory variables and insufficient data for their model.</p><p>The resulting coefficients from regression analyses with min-max scaled variables are interpreted as the change in the response variable, with an increase in the explanatory variable from the lowest to highest observed value.</p><p>Here, variables are transformed to quantify the difference of each observation from the mean in terms of standard deviations, whereby the data distribution is scaled to a mean of zero and standard deviation of one. Regression coefficients from z-standardized data represent the estimated change in the response variable per one unit increase in explanatory variables, where one unit represents one standard deviation in the z-standardized data.</p><p>Both transformations improve direct comparison between regression coefficients for variables measured on different scales, but differences in the algebraic transformation have considerable impact on conclusions. To demonstrate that Dickie et al. (<span>2024</span>)'s conclusions are sensitive to transformations, we z-standardized their original data and re-ran their global model examining effects of several variables on white-tailed deer density. Using min-max scaling, the authors originally reported negative statistically significant effects of Climate Dimension 1 (<i>β</i> = −6.794 ± 2.523, <i>p</i> < .007) and negative but insignificant effect of % Habitat Alteration (<i>β</i> = −0.328 ± 2.060, P = 0.873) (their Table 2). However, when using z-standardized data, we found a different conclusion of approximately equal magnitude but opposing effects of Climate Dimension 1 (<i>β</i> = −0.907 ± 0.286, <i>p</i> < .002) and % Habitat Alteration (<i>β</i> = 0.926 ± 0.307, <i>p</i> < .003, Figure 1).</p><p>Importantly, Dickie et al. (<span>2024</span>) describe a large dataset: “<i>…300 remote cameras across 12 replicated 50km</i><sup><i>2</i></sup> <i>landscapes over 5 years</i>.” and, “<i>Camera traps operated between 53,506 and 96,096 trap days from 2017 to 2021</i>”. However, the authors collapsed their response variable into a single mean density estimate for each camera cluster per year. The global model containing seven fixed effects (intercept, three “direct” effects, and three interactions), and two random effects, relies on only 53 data points. With a generous guideline of a minimum of 10 observations per treatment (Bolker et al., <span>2009</span>), their model has insufficient data to robustly estimate parameters of interest. This is likely the cause of large standard errors reported in their Table 2. The authors do admit estimates are imprecise, but do little to temper interpretations on climate vs. habitat alteration. We therefore constructed reduced models, which corroborate approximately equal but opposite effects of climate and habitat alteration on predicted deer density (Supplemental Materials S1).</p><p>Our reanalysis suggests both climate and habitat alteration drive density of white-tailed deer, but more extensive data across their Canadian range is required to estimate this properly. Parsing out the relative and interacting roles of these variables will be notoriously difficult at the landscape scale. Our reanalysis illustrates inherent risk of drawing potentially erroneous conclusions following different analytical decisions that produce contrasting results (Gould et al. <span>2023</span>). This is particularly problematic when conclusions are presented in social debates with direct impacts on conservation decisions by government authorities, as in woodland caribou conservation (CBC, <span>2024</span>). Given effective conservation actions depends on high-quality evidence, claims of which variables do and do not affect density of white-tailed deer in the boreal forest based on specific analytical decisions must be scrutinized.</p><p><b>Andrew Barnas:</b> Conceptualization; formal analysis; writing – original draft; writing – review and editing. <b>Brad Anholt:</b> Conceptualization; writing – original draft; writing – review and editing. <b>A. Cole Burton:</b> Conceptualization; writing – original draft; writing – review and editing. <b>Kathleen Carroll:</b> Conceptualization; writing – original draft; writing – review and editing. <b>Steeve D. Côté:</b> Conceptualization; writing – original draft; writing – review and editing. <b>Marco Festa-Bianchet:</b> Conceptualization; writing – original draft; writing – review and editing. <b>John Fryxell:</b> Conceptualization; writing – original draft; writing – review and editing. <b>Martin-Hugues St-Laurent:</b> Conceptualization; writing – original draft; writing – review and editing. <b>Jason T. Fisher:</b> Conceptualization; formal analysis; writing – original draft; writing – review and editing.</p><p>We have no conflict of interest to declare.</p>\",\"PeriodicalId\":175,\"journal\":{\"name\":\"Global Change Biology\",\"volume\":\"30 9\",\"pages\":\"\"},\"PeriodicalIF\":10.8000,\"publicationDate\":\"2024-09-13\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://onlinelibrary.wiley.com/doi/epdf/10.1111/gcb.17498\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"Global Change Biology\",\"FirstCategoryId\":\"93\",\"ListUrlMain\":\"https://onlinelibrary.wiley.com/doi/10.1111/gcb.17498\",\"RegionNum\":1,\"RegionCategory\":\"环境科学与生态学\",\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"Q1\",\"JCRName\":\"BIODIVERSITY CONSERVATION\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"Global Change Biology","FirstCategoryId":"93","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1111/gcb.17498","RegionNum":1,"RegionCategory":"环境科学与生态学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"BIODIVERSITY CONSERVATION","Score":null,"Total":0}
The influence of habitat alteration on density of invading white-tailed deer should not be discounted
White-tailed deer (Odocoileus virginianus) range expansion into boreal forests facilitates wolf (Canis lupus) population growth in many parts of Canada and is associated with caribou (Rangifer tarandus caribou) declines (Latham et al., 2011). Several works across Canada have demonstrated anthropogenic landscape change subsidizes forage available to white-tailed deer (Darlington et al., 2022; Fisher et al., 2020). However, recently Dickie et al. (2024) suggested instead climate is the primary driver of spatial variation in white-tailed deer density in Canada's boreal forest. These findings garnered significant media attention (CBC, 2024; NPR, 2024) with direct implications for conservation actions, specifically habitat protection and restoration efforts. We generally agree with the authors' conclusion that spatial variation in winter severity impacts deer population density, but we contend their conclusion on negligible impact of habitat alteration was compromised by their method of transforming explanatory variables and insufficient data for their model.
The resulting coefficients from regression analyses with min-max scaled variables are interpreted as the change in the response variable, with an increase in the explanatory variable from the lowest to highest observed value.
Here, variables are transformed to quantify the difference of each observation from the mean in terms of standard deviations, whereby the data distribution is scaled to a mean of zero and standard deviation of one. Regression coefficients from z-standardized data represent the estimated change in the response variable per one unit increase in explanatory variables, where one unit represents one standard deviation in the z-standardized data.
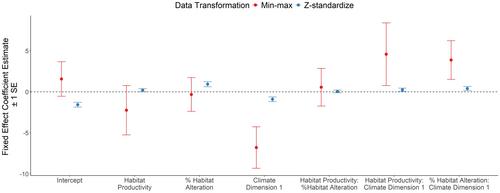
Both transformations improve direct comparison between regression coefficients for variables measured on different scales, but differences in the algebraic transformation have considerable impact on conclusions. To demonstrate that Dickie et al. (2024)'s conclusions are sensitive to transformations, we z-standardized their original data and re-ran their global model examining effects of several variables on white-tailed deer density. Using min-max scaling, the authors originally reported negative statistically significant effects of Climate Dimension 1 (β = −6.794 ± 2.523, p < .007) and negative but insignificant effect of % Habitat Alteration (β = −0.328 ± 2.060, P = 0.873) (their Table 2). However, when using z-standardized data, we found a different conclusion of approximately equal magnitude but opposing effects of Climate Dimension 1 (β = −0.907 ± 0.286, p < .002) and % Habitat Alteration (β = 0.926 ± 0.307, p < .003, Figure 1).
Importantly, Dickie et al. (2024) describe a large dataset: “…300 remote cameras across 12 replicated 50km2landscapes over 5 years.” and, “Camera traps operated between 53,506 and 96,096 trap days from 2017 to 2021”. However, the authors collapsed their response variable into a single mean density estimate for each camera cluster per year. The global model containing seven fixed effects (intercept, three “direct” effects, and three interactions), and two random effects, relies on only 53 data points. With a generous guideline of a minimum of 10 observations per treatment (Bolker et al., 2009), their model has insufficient data to robustly estimate parameters of interest. This is likely the cause of large standard errors reported in their Table 2. The authors do admit estimates are imprecise, but do little to temper interpretations on climate vs. habitat alteration. We therefore constructed reduced models, which corroborate approximately equal but opposite effects of climate and habitat alteration on predicted deer density (Supplemental Materials S1).
Our reanalysis suggests both climate and habitat alteration drive density of white-tailed deer, but more extensive data across their Canadian range is required to estimate this properly. Parsing out the relative and interacting roles of these variables will be notoriously difficult at the landscape scale. Our reanalysis illustrates inherent risk of drawing potentially erroneous conclusions following different analytical decisions that produce contrasting results (Gould et al. 2023). This is particularly problematic when conclusions are presented in social debates with direct impacts on conservation decisions by government authorities, as in woodland caribou conservation (CBC, 2024). Given effective conservation actions depends on high-quality evidence, claims of which variables do and do not affect density of white-tailed deer in the boreal forest based on specific analytical decisions must be scrutinized.
Andrew Barnas: Conceptualization; formal analysis; writing – original draft; writing – review and editing. Brad Anholt: Conceptualization; writing – original draft; writing – review and editing. A. Cole Burton: Conceptualization; writing – original draft; writing – review and editing. Kathleen Carroll: Conceptualization; writing – original draft; writing – review and editing. Steeve D. Côté: Conceptualization; writing – original draft; writing – review and editing. Marco Festa-Bianchet: Conceptualization; writing – original draft; writing – review and editing. John Fryxell: Conceptualization; writing – original draft; writing – review and editing. Martin-Hugues St-Laurent: Conceptualization; writing – original draft; writing – review and editing. Jason T. Fisher: Conceptualization; formal analysis; writing – original draft; writing – review and editing.
期刊介绍:
Global Change Biology is an environmental change journal committed to shaping the future and addressing the world's most pressing challenges, including sustainability, climate change, environmental protection, food and water safety, and global health.
Dedicated to fostering a profound understanding of the impacts of global change on biological systems and offering innovative solutions, the journal publishes a diverse range of content, including primary research articles, technical advances, research reviews, reports, opinions, perspectives, commentaries, and letters. Starting with the 2024 volume, Global Change Biology will transition to an online-only format, enhancing accessibility and contributing to the evolution of scholarly communication.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


