{"title":"氧化猝灭导致水稻自交系乌比什体损失和雄性不育。","authors":"Chuanlin Shi, Shaohua Yang, Yan Cui, Zhan Xu, Bin Zhang, Mingliang Guo, Yiwang Zhu, Ying Yang, Feng Wang, Huaqing Liu, Yu Zhang, Qian Qian, Lianguang Shang","doi":"10.1111/nph.20023","DOIUrl":null,"url":null,"abstract":"<p>The anther is a critical male organ for plant reproduction in flowering plants. In rice (<i>Oryza sativa</i> L.), anthers consist of four somatic cell layers named epidermis, endothecium, middle layer and tapetum, which surround the reproductive cells (Zhang <i>et al</i>., <span>2011</span>). The rice anther is of particular interest for crop breeding researchers because the male sterile line, mostly resulting from anther developmental defect, is the core component for hybrid rice breeding. Ubisch bodies, the unique U-shaped secretory structures distributed in the inner surface of tapetum, mediate exporting tapetum-produced sugars, lipids, proteins and sporopollenin precursors to facilitate microspore development. Reactive oxygen species (ROS), including H<sub>2</sub>O<sub>2</sub> and superoxide anion (O<sub>2</sub><sup>•−</sup>) as important oxidizing agents in living cells, have been reported to trigger tapetal programmed cell death (PCD) and thus male sterility in rice (Hu <i>et al</i>., <span>2011</span>; Luo <i>et al</i>., <span>2013</span>). However, the regulation of ROS homeostasis in tapetum development, Ubisch body formation and its impact on male fertility remain unclear.</p><p>To gain insight into the genetic regulation of male sterility in rice, we identified a mutant named <i>no ubisch body and pollen grain 1</i> (<i>nup1</i>) from a tissue culture-induced mutant library in the <i>indica</i> cv MingHui86 (MH86). At maturity, the <i>nup1</i> mutant exhibited a slightly enclosed panicle, and smaller and paler yellow anthers with no pollen grain and seed production compared with the wild-type (WT) MH86, while plant architecture, female organ and spikelet showed no obvious differences (Fig. 1a–e, Supporting Information Fig. S1), indicating that the <i>nup1</i> mutant is male sterility. Upon pollination of the <i>nup1</i> mutant with the pollen grains of WT, all the F<sub>1</sub> plants exhibited normal fertility and the F<sub>2</sub> plants showed a segregation of 249 fertile and 78 sterile plants (3 : 1, <i>χ</i><sup>2</sup> = 0.26, <i>P</i> > 0.05), demonstrating that this male sterile phenotype is controlled by a single recessive gene. To investigate the cytological defects of the <i>nup1</i> mutant, anthers were analyzed through semi-thin sections based on anther developmental stages as described previously (Zhang <i>et al</i>., <span>2011</span>). We found that no differences were visible between the <i>nup1</i> mutant and WT during Stage 7 (S7) to Stage 9 (S9) (Fig. S2a). Notably, the <i>nup1</i> mutant exhibited obviously shrinking tapetal cells and swollen microspores at S10 and S11, but the swollen microspores rapidly shriveled and crushed at late S11 and S12 (Figs 1f,g, S2b), resulting in no pollen grain production compared with the WT which underwent the regular anther developments.</p><p>Transmission electron microscopy (TEM) analysis was performed as described in Methods S1 to observe more detailed structure changes in <i>nup1</i> anthers. At Stage 8b (S8b), both the <i>nup1</i> mutant and WT underwent meiosis normally to produce the tetrads, and their tapetal cells were covered by a thin layer of cell wall with no Ubisch body formation (Fig. S3a–f). At Stage 9 (S9), WT tapetal cells started to produce Ubisch bodies, whereas in the <i>nup1</i> mutant, the Ubisch bodies did not yet emerged on tapetal cells (Fig. S3g–l). At S10, WT tapetal cells were more condensed and formed a large number of characteristic Ubisch bodies on their inner surface, and the pollen walls formed well-organized structures including nexine, bacula and tectum (Figs 1h, S4a). However, interestingly, no Ubisch bodies emerged on the surface of the <i>nup1</i> tapetum, and the bacula was missing accompanied by narrowed nexine and tectum in the <i>nup1</i> mutant (Figs 1i, S4b). During S11 to S12, still no Ubisch bodies were observed, and all pollen grains collapsed with tectum and nexine appearing as a much looser two-layer structure in the <i>nup1</i> mutant (Fig. S4c–n). These results indicated that Ubisch body formation during anther development was completely absent in <i>nup1</i> mutant anthers. Moreover, scanning electron microscopy (SEM) analysis of S12 anthers showed that the cuticle of WT anther displayed the typical spaghetti-like pattern, whereas the <i>nup1</i> anther surface lacked the cuticle structure and appeared nearly smooth (Figs 1j,k, S5a,b). In agreement with TEM observations, abundant Ubisch bodies were observed with crystal-like shapes on the locular side of WT anthers, but these structures were completely absent in <i>nup1</i> anthers (Fig. 1l,m), confirming the loss of Ubisch body in the <i>nup1</i> mutant. This serious absence of Ubisch body during anther development has not been thoroughly explored in rice. Given these unique characteristics, we thus named this mutant <i>no ubisch body and pollen grain 1</i> (<i>nup1</i>). Taken together, these results indicate that <i>NUP1</i> is essentially required for the development of tapetum, anther cuticle, pollen wall and, particularly, the formation of Ubisch bodies.</p><p>To test whether the disappearance of the Ubisch body was due to the possible premature degradation of the tapetum in the <i>nup1</i> mutant, we performed the terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling (TUNEL) assay to detect the timely PCD of the tapetum between WT and the <i>nup1</i> mutant. In both WT and <i>nup1</i> anthers, the positive TUNEL signals were clearly visualized at S7–S9 and declined during S10–S11, and there were no obvious differences in TUNEL signals between WT and the <i>nup1</i> mutant during the anther developmental stages (Fig. S6). Although the cytological analysis showed that the <i>nup1</i> mutant displayed shrinking tapetal cells at S10 and S11, we did not observe premature degradation of tapetal cells in <i>nup1</i> mutant anthers compared with WT. These results indicated that loss-of-function of <i>NUP1</i> did not influence PCD of tapetal cells, but contributed to the loss of Ubisch body formation, which resulted in male sterility in the <i>nup1</i> mutant.</p><p>To clone the <i>NUP1</i> gene, we generated the F<sub>2</sub> population from a cross between the <i>nup1</i> mutant and <i>indica</i> cv 9311. Using 50 F<sub>2</sub> individuals with the <i>nup1</i> mutant phenotype for bulked segregant analysis by deep sequencing (BSA-seq), the <i>NUP1</i> locus was limited to a 10.17-Mb region on chromosome 8 (Fig. 1n). By a map-based cloning with designed molecular markers (Table S1), we further narrowed the <i>NUP1</i> locus to a 121-kb region (Fig. 1o). Whole-genome re-sequencing analysis between WT and the <i>nup1</i> mutant revealed a single-nucleotide C-to-A substitution in gene coding region within the locus, and this substitution was occurred in the fourth exon of LOC_Os08g20730 (Fig. 1p), resulting in a premature stop codon. Therefore, we identified LOC_Os08g20730 which encodes a class III peroxidase as the candidate gene. To validate this, we developed <i>NUP1</i> knockout transgenic plants in WT MH86 using the CRISPR/Cas9 approach. Five independent knockout (KO) lines were confirmed to have shifted open reading frames in the first exon (Figs 1p, S7a) and showed smaller and paler anthers with no pollen grains and seed production (Figs 1q,r, S4b–f), identical to the <i>nup1</i> mutant phenotype. Therefore, we concluded that the <i>NUP1</i> gene was LOC_Os08g20730.</p><p>To determine the expression pattern of <i>NUP1</i>, we performed reverse transcription quantitative polymerase chain reaction and found that <i>NUP1</i> was preferentially expressed in the anthers during developmental S6 to S12 and lowly expressed in vegetative organs (Fig. 1s). The RNA <i>in situ</i> hybridization of <i>NUP1</i> in WT showed that <i>NUP1</i> transcripts were mainly accumulated in tapetal cells and slightly distributed in other cell layers of anther wall and microspores at different developmental stages in anthers (Fig. S8). Furthermore, we generated <i>NUP1-GFP</i> transgenic plants using its native promoter (<i>pNUP1::NUP1-GFP</i>) and observed that NUP1-GFP fusion proteins were substantially localized in all layers of anther wall including tapetum across S7 to S9 (Fig. S9). The rare GFP signal in microspores may be due to a low level of fusion proteins to be observed. Collectively, this expression pattern of <i>NUP1</i> in anthers confirmed that <i>NUP1</i> played an essential role in tapetum and microspore development. To determine the subcellular localization of NUP1, we transiently expressed a construct encoding NUP1-GFP fusion protein together with a construct encoding OsH2B-mCherry (histone OsH2B-mCherry fusion, as a nuclear marker) in <i>Nicotiana benthamiana</i> leaves. The fluorescent signals of NUP1-GFP were dominantly localized in the cytoplasm, excluding with OsH2B-mCherry signals (Fig. 1t). The transfection of rice protoplasts with construct encoding NUP1-YFP fusion further confirmed the cytoplasm-localization of NUP1 protein (Fig. S10).</p><p>Class III peroxidases are a large family of plant-specific heme-iron–dependent peroxidases that use hydrogen peroxide as the electron acceptor to catalyze a number of oxidative reactions (Shigeto & Tsutsumi, <span>2016</span>). There are 73 peroxidase genes in <i>Arabidopsis thaliana</i> and 138 genes in rice. These genes are involved in a range of functions, such as lignification, cell elongation and stress defense (Shigeto & Tsutsumi, <span>2016</span>). Phylogenetic analysis showed that <i>NUP1</i> was highly conserved among different plant species (Fig. S11). Notably, its closely related homolog in <i>Arabidopsis</i>, <i>PRX40</i>, has been found to function redundantly with <i>PRX9</i> to maintain tapetum and microspore development (Jacobowitz <i>et al</i>., <span>2019</span>). Amino acid alignment indicated that the enzyme active sites and heme-ligand binding sites were highly conserved among NUP1, PRX40 and other homologs (Fig. S12). However, even though <i>PRX9</i> and <i>PRX14</i>, two homologs of rice <i>NUP1</i> in <i>Arabidopsis</i> have been found to be responsible for <i>Arabidopsis</i> anther development, there are several significant differences in their functions between <i>Arabidopsis</i> and rice: (1) single mutants of either <i>prx9</i> or <i>prx40</i> displayed fully fertile, and only <i>prx9 prx40</i> double-mutant plants were completely sterile in <i>Arabidopsis</i>, whereas single mutant of <i>nup1</i> exhibits male sterility in rice; (2) tapetal cells of <i>prx9 prx40</i> double mutants were swollen and invaded into the locular space, while tapetal cells of <i>nup1</i> in rice were, to some extent, shriveled with absolutely no Ubisch body formation. Therefore, <i>NUP1</i> and its homologs, <i>PRX9</i> and <i>PRX14</i>, exhibit some functional diversity in anther development between monocot rice and dicot <i>Arabidopsis</i>.</p><p>To confirm whether NUP1-mediated ROS scavenging as a peroxidase, we performed nitroblue tetrazolium (NBT) staining to determine the difference in ROS accumulation during anther development between WT and the <i>nup1</i> mutant. In WT, the ROS signals were clearly detectable during S6 to S9, declined at S10, were almost undetectable at S11, and were re-detectable at S12 (Fig. 2a), which were consistent with previous study (Zhou <i>et al</i>., <span>2024</span>). During S10 to S11, tapetal cells become degenerated and form Ubisch bodies abundantly, and microspores undergo mitosis, accumulate starch and synthesize the pollen walls using nutrition delivered by Ubisch bodies (Zhang <i>et al</i>., <span>2011</span>). However, in the <i>nup1</i> mutant and knockout line KO#1, increased ROS signals were observed at S9, and strong ROS signals were detected at S10 to S11 compared with WT (Figs 2b, S13a,b). Furthermore, we directly measured the contents of H<sub>2</sub>O<sub>2</sub> and malondialdehyde (MDA), which is an indicator of local ROS production and participant in peroxidation of fatty acids, in anthers at various stages. In agreement with the NBT staining, the contents of H<sub>2</sub>O<sub>2</sub> and MDA were comparable in anthers between WT and the <i>nup1</i> mutant during S6 to S9, whereas they were significantly increased in <i>nup1</i> anthers compared with WT at S10 and S11 (Figs 2c, S13c), suggesting a ROS burst in <i>nup1</i> anthers at the later developmental stages. Consistently, a recent study reported that under high temperature excessive ROS in anthers at S10 to S11 in <i>ostms19</i>, <i>tms5</i>, <i>ostms15</i> and <i>ostms18</i> is responsible for their high temperature-sensitive male sterility (Zhou <i>et al</i>., <span>2024</span>). Taking together, these results indicate that the timing of ROS burst in the <i>nup1</i> mutant corresponds to the defect in <i>nup1</i> anthers and suggests that <i>NUP1</i> is involved in ROS scavenging, which is important for Ubisch body formation and microspore development during S10 to S11.</p><p>To investigate the impacts of ROS burst on gene expression in <i>nup1</i> anther, we conducted anther transcriptome profiling (Fig. S14a). Differentially expressed genes (DEGs) analysis (FDR < 0.01, FC > 2) showed that thousands of genes were significantly changed in <i>nup1</i> anthers compared with WT at S10 and S11 (Fig. S14b,c; Tables S2, S3). We identified 525 upregulated and 726 downregulated DEGs overlapped between anthers at S10 and S11 (Fig. 2d). Gene Ontology (GO) analysis showed that obsolete oxidation–reduction process was the most significantly enriched GO term for DEGs at S10, S11 and the overlapped DEGs at both stages (Fig. 2e), consistent with the ROS burst in the <i>nup1</i> mutant. Furthermore, the carbohydrate metabolic process, the lipid metabolic process and transmembrane transport were among the top-listed significantly enriched GO terms at both S10 and S11 (Fig. 2e). Indeed, we identified and confirmed that many known genes associated with oxidation–reduction, carbohydrate and lipid metabolisms were significantly up- and downregulated in <i>nup1</i> anthers compared with WT at both stages (Figs 2d, S14d, S15). Carbohydrate and lipid metabolisms have been widely demonstrated to play critical roles in tapetum development and pollen formation (Wan <i>et al</i>., <span>2020</span>; Liu <i>et al</i>., <span>2021</span>). These results indicated that ROS burst in <i>nup1</i> anthers resulted in defective oxidation–reduction processes and carbohydrate and lipid metabolisms.</p><p>To confirm the dysregulated metabolism, we performed mass spectrometry to identify the metabolites in S11 anthers of WT and the <i>nup1</i> mutant (Fig. S16a). A total of 1963 metabolites including flavonoids, amino acids and derivatives, phenolic acids, lipids, alkaloids, organic acids, lignans and coumarins, and others were identified (Fig. S16b). Differential metabolites analysis showed that 671 compounds were significantly changed in <i>nup1</i> anthers compared with WT at S11, of which flavonoids (32.34%), lipids (15.80%) and phenolic acids (11.77%) accounted for the top three dysregulated metabolites (Fig. S16c,d; Table S4). In line with the significant GO term related to lipid metabolism, lipid metabolites were predominantly increased in <i>nup1</i> anthers compared with WT (Fig. 2g). Notably, flavonoids, the antioxidants that mediate direct scavenging of ROS in plants (Agati <i>et al</i>., <span>2012</span>), were significantly reduced in <i>nup1</i> anthers (Fig. S16e), This reduction may be attributed to ROS consumption. Moreover, KEGG pathway analysis showed that two lipids, linoleic acid and arachidonic acid metabolism, as well as phenylpropanoid and flavonoid biosynthesis were the significantly enriched terms (Figs S16f, S17). These findings further confirm the dysregulated lipid and oxidation–reduction metabolisms in <i>nup1</i> anthers.</p><p>In summary, we identified the <i>nup1</i> mutant in rice that exhibited specific male sterility due to the complete absence of Ubisch body formation and pollen grain production. We cloned the causal gene and demonstrated that <i>NUP1</i>, encoding a class III peroxidase predominantly expressed in the anther wall, mediated ROS scavenging. The loss-of-function of <i>NUP1</i> resulted in ROS burst in anthers, which disrupted the oxidation–reduction process, carbohydrate and lipid metabolisms, and loss of tapetal Ubisch body, thereby resulting in male sterility in rice (Fig. 2h). Our study expands the understanding of the molecular mechanisms by which ROS homeostasis regulates cell metabolism, tapetal Ubisch body formation and male fertility in plants. Recently, the third-generation hybrid rice technology using a recessive nuclear male sterile line has been successfully demonstrated to develop genetic systems for hybrid rice (Chang <i>et al</i>., <span>2016</span>; Liao <i>et al</i>., <span>2021</span>). Since the loss of <i>NUP1</i> resulted in completely absent Ubisch body formation and no pollen grain production without changing female fertility and plant growth, <i>NUP1</i> is a potentially useful male sterility gene in third-generation hybrid rice breeding in the future.</p><p>None declared.</p><p>LS, QQ, Y Zhang and HL conceived and designed this research. CS, SY and YC performed most of the experiments. ZX, BZ, MG, Y Zhu and YY planted the materials and performed the gene cloning. CS and YC performed data analysis. CS and SY wrote the manuscript. LS, QQ, Y Zhang, HL and FW provided guidance in writing the manuscript. All authors contributed to editing the manuscript. CS, SY, YC and ZX contributed equally to this work.</p>","PeriodicalId":214,"journal":{"name":"New Phytologist","volume":null,"pages":null},"PeriodicalIF":8.3000,"publicationDate":"2024-08-01","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1111/nph.20023","citationCount":"0","resultStr":"{\"title\":\"Oxidative burst causes loss of tapetal Ubisch body and male sterility in rice\",\"authors\":\"Chuanlin Shi, Shaohua Yang, Yan Cui, Zhan Xu, Bin Zhang, Mingliang Guo, Yiwang Zhu, Ying Yang, Feng Wang, Huaqing Liu, Yu Zhang, Qian Qian, Lianguang Shang\",\"doi\":\"10.1111/nph.20023\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p>The anther is a critical male organ for plant reproduction in flowering plants. In rice (<i>Oryza sativa</i> L.), anthers consist of four somatic cell layers named epidermis, endothecium, middle layer and tapetum, which surround the reproductive cells (Zhang <i>et al</i>., <span>2011</span>). The rice anther is of particular interest for crop breeding researchers because the male sterile line, mostly resulting from anther developmental defect, is the core component for hybrid rice breeding. Ubisch bodies, the unique U-shaped secretory structures distributed in the inner surface of tapetum, mediate exporting tapetum-produced sugars, lipids, proteins and sporopollenin precursors to facilitate microspore development. Reactive oxygen species (ROS), including H<sub>2</sub>O<sub>2</sub> and superoxide anion (O<sub>2</sub><sup>•−</sup>) as important oxidizing agents in living cells, have been reported to trigger tapetal programmed cell death (PCD) and thus male sterility in rice (Hu <i>et al</i>., <span>2011</span>; Luo <i>et al</i>., <span>2013</span>). However, the regulation of ROS homeostasis in tapetum development, Ubisch body formation and its impact on male fertility remain unclear.</p><p>To gain insight into the genetic regulation of male sterility in rice, we identified a mutant named <i>no ubisch body and pollen grain 1</i> (<i>nup1</i>) from a tissue culture-induced mutant library in the <i>indica</i> cv MingHui86 (MH86). At maturity, the <i>nup1</i> mutant exhibited a slightly enclosed panicle, and smaller and paler yellow anthers with no pollen grain and seed production compared with the wild-type (WT) MH86, while plant architecture, female organ and spikelet showed no obvious differences (Fig. 1a–e, Supporting Information Fig. S1), indicating that the <i>nup1</i> mutant is male sterility. Upon pollination of the <i>nup1</i> mutant with the pollen grains of WT, all the F<sub>1</sub> plants exhibited normal fertility and the F<sub>2</sub> plants showed a segregation of 249 fertile and 78 sterile plants (3 : 1, <i>χ</i><sup>2</sup> = 0.26, <i>P</i> > 0.05), demonstrating that this male sterile phenotype is controlled by a single recessive gene. To investigate the cytological defects of the <i>nup1</i> mutant, anthers were analyzed through semi-thin sections based on anther developmental stages as described previously (Zhang <i>et al</i>., <span>2011</span>). We found that no differences were visible between the <i>nup1</i> mutant and WT during Stage 7 (S7) to Stage 9 (S9) (Fig. S2a). Notably, the <i>nup1</i> mutant exhibited obviously shrinking tapetal cells and swollen microspores at S10 and S11, but the swollen microspores rapidly shriveled and crushed at late S11 and S12 (Figs 1f,g, S2b), resulting in no pollen grain production compared with the WT which underwent the regular anther developments.</p><p>Transmission electron microscopy (TEM) analysis was performed as described in Methods S1 to observe more detailed structure changes in <i>nup1</i> anthers. At Stage 8b (S8b), both the <i>nup1</i> mutant and WT underwent meiosis normally to produce the tetrads, and their tapetal cells were covered by a thin layer of cell wall with no Ubisch body formation (Fig. S3a–f). At Stage 9 (S9), WT tapetal cells started to produce Ubisch bodies, whereas in the <i>nup1</i> mutant, the Ubisch bodies did not yet emerged on tapetal cells (Fig. S3g–l). At S10, WT tapetal cells were more condensed and formed a large number of characteristic Ubisch bodies on their inner surface, and the pollen walls formed well-organized structures including nexine, bacula and tectum (Figs 1h, S4a). However, interestingly, no Ubisch bodies emerged on the surface of the <i>nup1</i> tapetum, and the bacula was missing accompanied by narrowed nexine and tectum in the <i>nup1</i> mutant (Figs 1i, S4b). During S11 to S12, still no Ubisch bodies were observed, and all pollen grains collapsed with tectum and nexine appearing as a much looser two-layer structure in the <i>nup1</i> mutant (Fig. S4c–n). These results indicated that Ubisch body formation during anther development was completely absent in <i>nup1</i> mutant anthers. Moreover, scanning electron microscopy (SEM) analysis of S12 anthers showed that the cuticle of WT anther displayed the typical spaghetti-like pattern, whereas the <i>nup1</i> anther surface lacked the cuticle structure and appeared nearly smooth (Figs 1j,k, S5a,b). In agreement with TEM observations, abundant Ubisch bodies were observed with crystal-like shapes on the locular side of WT anthers, but these structures were completely absent in <i>nup1</i> anthers (Fig. 1l,m), confirming the loss of Ubisch body in the <i>nup1</i> mutant. This serious absence of Ubisch body during anther development has not been thoroughly explored in rice. Given these unique characteristics, we thus named this mutant <i>no ubisch body and pollen grain 1</i> (<i>nup1</i>). Taken together, these results indicate that <i>NUP1</i> is essentially required for the development of tapetum, anther cuticle, pollen wall and, particularly, the formation of Ubisch bodies.</p><p>To test whether the disappearance of the Ubisch body was due to the possible premature degradation of the tapetum in the <i>nup1</i> mutant, we performed the terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling (TUNEL) assay to detect the timely PCD of the tapetum between WT and the <i>nup1</i> mutant. In both WT and <i>nup1</i> anthers, the positive TUNEL signals were clearly visualized at S7–S9 and declined during S10–S11, and there were no obvious differences in TUNEL signals between WT and the <i>nup1</i> mutant during the anther developmental stages (Fig. S6). Although the cytological analysis showed that the <i>nup1</i> mutant displayed shrinking tapetal cells at S10 and S11, we did not observe premature degradation of tapetal cells in <i>nup1</i> mutant anthers compared with WT. These results indicated that loss-of-function of <i>NUP1</i> did not influence PCD of tapetal cells, but contributed to the loss of Ubisch body formation, which resulted in male sterility in the <i>nup1</i> mutant.</p><p>To clone the <i>NUP1</i> gene, we generated the F<sub>2</sub> population from a cross between the <i>nup1</i> mutant and <i>indica</i> cv 9311. Using 50 F<sub>2</sub> individuals with the <i>nup1</i> mutant phenotype for bulked segregant analysis by deep sequencing (BSA-seq), the <i>NUP1</i> locus was limited to a 10.17-Mb region on chromosome 8 (Fig. 1n). By a map-based cloning with designed molecular markers (Table S1), we further narrowed the <i>NUP1</i> locus to a 121-kb region (Fig. 1o). Whole-genome re-sequencing analysis between WT and the <i>nup1</i> mutant revealed a single-nucleotide C-to-A substitution in gene coding region within the locus, and this substitution was occurred in the fourth exon of LOC_Os08g20730 (Fig. 1p), resulting in a premature stop codon. Therefore, we identified LOC_Os08g20730 which encodes a class III peroxidase as the candidate gene. To validate this, we developed <i>NUP1</i> knockout transgenic plants in WT MH86 using the CRISPR/Cas9 approach. Five independent knockout (KO) lines were confirmed to have shifted open reading frames in the first exon (Figs 1p, S7a) and showed smaller and paler anthers with no pollen grains and seed production (Figs 1q,r, S4b–f), identical to the <i>nup1</i> mutant phenotype. Therefore, we concluded that the <i>NUP1</i> gene was LOC_Os08g20730.</p><p>To determine the expression pattern of <i>NUP1</i>, we performed reverse transcription quantitative polymerase chain reaction and found that <i>NUP1</i> was preferentially expressed in the anthers during developmental S6 to S12 and lowly expressed in vegetative organs (Fig. 1s). The RNA <i>in situ</i> hybridization of <i>NUP1</i> in WT showed that <i>NUP1</i> transcripts were mainly accumulated in tapetal cells and slightly distributed in other cell layers of anther wall and microspores at different developmental stages in anthers (Fig. S8). Furthermore, we generated <i>NUP1-GFP</i> transgenic plants using its native promoter (<i>pNUP1::NUP1-GFP</i>) and observed that NUP1-GFP fusion proteins were substantially localized in all layers of anther wall including tapetum across S7 to S9 (Fig. S9). The rare GFP signal in microspores may be due to a low level of fusion proteins to be observed. Collectively, this expression pattern of <i>NUP1</i> in anthers confirmed that <i>NUP1</i> played an essential role in tapetum and microspore development. To determine the subcellular localization of NUP1, we transiently expressed a construct encoding NUP1-GFP fusion protein together with a construct encoding OsH2B-mCherry (histone OsH2B-mCherry fusion, as a nuclear marker) in <i>Nicotiana benthamiana</i> leaves. The fluorescent signals of NUP1-GFP were dominantly localized in the cytoplasm, excluding with OsH2B-mCherry signals (Fig. 1t). The transfection of rice protoplasts with construct encoding NUP1-YFP fusion further confirmed the cytoplasm-localization of NUP1 protein (Fig. S10).</p><p>Class III peroxidases are a large family of plant-specific heme-iron–dependent peroxidases that use hydrogen peroxide as the electron acceptor to catalyze a number of oxidative reactions (Shigeto & Tsutsumi, <span>2016</span>). There are 73 peroxidase genes in <i>Arabidopsis thaliana</i> and 138 genes in rice. These genes are involved in a range of functions, such as lignification, cell elongation and stress defense (Shigeto & Tsutsumi, <span>2016</span>). Phylogenetic analysis showed that <i>NUP1</i> was highly conserved among different plant species (Fig. S11). Notably, its closely related homolog in <i>Arabidopsis</i>, <i>PRX40</i>, has been found to function redundantly with <i>PRX9</i> to maintain tapetum and microspore development (Jacobowitz <i>et al</i>., <span>2019</span>). Amino acid alignment indicated that the enzyme active sites and heme-ligand binding sites were highly conserved among NUP1, PRX40 and other homologs (Fig. S12). However, even though <i>PRX9</i> and <i>PRX14</i>, two homologs of rice <i>NUP1</i> in <i>Arabidopsis</i> have been found to be responsible for <i>Arabidopsis</i> anther development, there are several significant differences in their functions between <i>Arabidopsis</i> and rice: (1) single mutants of either <i>prx9</i> or <i>prx40</i> displayed fully fertile, and only <i>prx9 prx40</i> double-mutant plants were completely sterile in <i>Arabidopsis</i>, whereas single mutant of <i>nup1</i> exhibits male sterility in rice; (2) tapetal cells of <i>prx9 prx40</i> double mutants were swollen and invaded into the locular space, while tapetal cells of <i>nup1</i> in rice were, to some extent, shriveled with absolutely no Ubisch body formation. Therefore, <i>NUP1</i> and its homologs, <i>PRX9</i> and <i>PRX14</i>, exhibit some functional diversity in anther development between monocot rice and dicot <i>Arabidopsis</i>.</p><p>To confirm whether NUP1-mediated ROS scavenging as a peroxidase, we performed nitroblue tetrazolium (NBT) staining to determine the difference in ROS accumulation during anther development between WT and the <i>nup1</i> mutant. In WT, the ROS signals were clearly detectable during S6 to S9, declined at S10, were almost undetectable at S11, and were re-detectable at S12 (Fig. 2a), which were consistent with previous study (Zhou <i>et al</i>., <span>2024</span>). During S10 to S11, tapetal cells become degenerated and form Ubisch bodies abundantly, and microspores undergo mitosis, accumulate starch and synthesize the pollen walls using nutrition delivered by Ubisch bodies (Zhang <i>et al</i>., <span>2011</span>). However, in the <i>nup1</i> mutant and knockout line KO#1, increased ROS signals were observed at S9, and strong ROS signals were detected at S10 to S11 compared with WT (Figs 2b, S13a,b). Furthermore, we directly measured the contents of H<sub>2</sub>O<sub>2</sub> and malondialdehyde (MDA), which is an indicator of local ROS production and participant in peroxidation of fatty acids, in anthers at various stages. In agreement with the NBT staining, the contents of H<sub>2</sub>O<sub>2</sub> and MDA were comparable in anthers between WT and the <i>nup1</i> mutant during S6 to S9, whereas they were significantly increased in <i>nup1</i> anthers compared with WT at S10 and S11 (Figs 2c, S13c), suggesting a ROS burst in <i>nup1</i> anthers at the later developmental stages. Consistently, a recent study reported that under high temperature excessive ROS in anthers at S10 to S11 in <i>ostms19</i>, <i>tms5</i>, <i>ostms15</i> and <i>ostms18</i> is responsible for their high temperature-sensitive male sterility (Zhou <i>et al</i>., <span>2024</span>). Taking together, these results indicate that the timing of ROS burst in the <i>nup1</i> mutant corresponds to the defect in <i>nup1</i> anthers and suggests that <i>NUP1</i> is involved in ROS scavenging, which is important for Ubisch body formation and microspore development during S10 to S11.</p><p>To investigate the impacts of ROS burst on gene expression in <i>nup1</i> anther, we conducted anther transcriptome profiling (Fig. S14a). Differentially expressed genes (DEGs) analysis (FDR < 0.01, FC > 2) showed that thousands of genes were significantly changed in <i>nup1</i> anthers compared with WT at S10 and S11 (Fig. S14b,c; Tables S2, S3). We identified 525 upregulated and 726 downregulated DEGs overlapped between anthers at S10 and S11 (Fig. 2d). Gene Ontology (GO) analysis showed that obsolete oxidation–reduction process was the most significantly enriched GO term for DEGs at S10, S11 and the overlapped DEGs at both stages (Fig. 2e), consistent with the ROS burst in the <i>nup1</i> mutant. Furthermore, the carbohydrate metabolic process, the lipid metabolic process and transmembrane transport were among the top-listed significantly enriched GO terms at both S10 and S11 (Fig. 2e). Indeed, we identified and confirmed that many known genes associated with oxidation–reduction, carbohydrate and lipid metabolisms were significantly up- and downregulated in <i>nup1</i> anthers compared with WT at both stages (Figs 2d, S14d, S15). Carbohydrate and lipid metabolisms have been widely demonstrated to play critical roles in tapetum development and pollen formation (Wan <i>et al</i>., <span>2020</span>; Liu <i>et al</i>., <span>2021</span>). These results indicated that ROS burst in <i>nup1</i> anthers resulted in defective oxidation–reduction processes and carbohydrate and lipid metabolisms.</p><p>To confirm the dysregulated metabolism, we performed mass spectrometry to identify the metabolites in S11 anthers of WT and the <i>nup1</i> mutant (Fig. S16a). A total of 1963 metabolites including flavonoids, amino acids and derivatives, phenolic acids, lipids, alkaloids, organic acids, lignans and coumarins, and others were identified (Fig. S16b). Differential metabolites analysis showed that 671 compounds were significantly changed in <i>nup1</i> anthers compared with WT at S11, of which flavonoids (32.34%), lipids (15.80%) and phenolic acids (11.77%) accounted for the top three dysregulated metabolites (Fig. S16c,d; Table S4). In line with the significant GO term related to lipid metabolism, lipid metabolites were predominantly increased in <i>nup1</i> anthers compared with WT (Fig. 2g). Notably, flavonoids, the antioxidants that mediate direct scavenging of ROS in plants (Agati <i>et al</i>., <span>2012</span>), were significantly reduced in <i>nup1</i> anthers (Fig. S16e), This reduction may be attributed to ROS consumption. Moreover, KEGG pathway analysis showed that two lipids, linoleic acid and arachidonic acid metabolism, as well as phenylpropanoid and flavonoid biosynthesis were the significantly enriched terms (Figs S16f, S17). These findings further confirm the dysregulated lipid and oxidation–reduction metabolisms in <i>nup1</i> anthers.</p><p>In summary, we identified the <i>nup1</i> mutant in rice that exhibited specific male sterility due to the complete absence of Ubisch body formation and pollen grain production. We cloned the causal gene and demonstrated that <i>NUP1</i>, encoding a class III peroxidase predominantly expressed in the anther wall, mediated ROS scavenging. The loss-of-function of <i>NUP1</i> resulted in ROS burst in anthers, which disrupted the oxidation–reduction process, carbohydrate and lipid metabolisms, and loss of tapetal Ubisch body, thereby resulting in male sterility in rice (Fig. 2h). Our study expands the understanding of the molecular mechanisms by which ROS homeostasis regulates cell metabolism, tapetal Ubisch body formation and male fertility in plants. Recently, the third-generation hybrid rice technology using a recessive nuclear male sterile line has been successfully demonstrated to develop genetic systems for hybrid rice (Chang <i>et al</i>., <span>2016</span>; Liao <i>et al</i>., <span>2021</span>). Since the loss of <i>NUP1</i> resulted in completely absent Ubisch body formation and no pollen grain production without changing female fertility and plant growth, <i>NUP1</i> is a potentially useful male sterility gene in third-generation hybrid rice breeding in the future.</p><p>None declared.</p><p>LS, QQ, Y Zhang and HL conceived and designed this research. CS, SY and YC performed most of the experiments. ZX, BZ, MG, Y Zhu and YY planted the materials and performed the gene cloning. CS and YC performed data analysis. CS and SY wrote the manuscript. LS, QQ, Y Zhang, HL and FW provided guidance in writing the manuscript. All authors contributed to editing the manuscript. CS, SY, YC and ZX contributed equally to this work.</p>\",\"PeriodicalId\":214,\"journal\":{\"name\":\"New Phytologist\",\"volume\":null,\"pages\":null},\"PeriodicalIF\":8.3000,\"publicationDate\":\"2024-08-01\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://onlinelibrary.wiley.com/doi/epdf/10.1111/nph.20023\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"New Phytologist\",\"FirstCategoryId\":\"99\",\"ListUrlMain\":\"https://onlinelibrary.wiley.com/doi/10.1111/nph.20023\",\"RegionNum\":1,\"RegionCategory\":\"生物学\",\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"Q1\",\"JCRName\":\"PLANT SCIENCES\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"New Phytologist","FirstCategoryId":"99","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1111/nph.20023","RegionNum":1,"RegionCategory":"生物学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"PLANT SCIENCES","Score":null,"Total":0}
Oxidative burst causes loss of tapetal Ubisch body and male sterility in rice
The anther is a critical male organ for plant reproduction in flowering plants. In rice (Oryza sativa L.), anthers consist of four somatic cell layers named epidermis, endothecium, middle layer and tapetum, which surround the reproductive cells (Zhang et al., 2011). The rice anther is of particular interest for crop breeding researchers because the male sterile line, mostly resulting from anther developmental defect, is the core component for hybrid rice breeding. Ubisch bodies, the unique U-shaped secretory structures distributed in the inner surface of tapetum, mediate exporting tapetum-produced sugars, lipids, proteins and sporopollenin precursors to facilitate microspore development. Reactive oxygen species (ROS), including H2O2 and superoxide anion (O2•−) as important oxidizing agents in living cells, have been reported to trigger tapetal programmed cell death (PCD) and thus male sterility in rice (Hu et al., 2011; Luo et al., 2013). However, the regulation of ROS homeostasis in tapetum development, Ubisch body formation and its impact on male fertility remain unclear.
To gain insight into the genetic regulation of male sterility in rice, we identified a mutant named no ubisch body and pollen grain 1 (nup1) from a tissue culture-induced mutant library in the indica cv MingHui86 (MH86). At maturity, the nup1 mutant exhibited a slightly enclosed panicle, and smaller and paler yellow anthers with no pollen grain and seed production compared with the wild-type (WT) MH86, while plant architecture, female organ and spikelet showed no obvious differences (Fig. 1a–e, Supporting Information Fig. S1), indicating that the nup1 mutant is male sterility. Upon pollination of the nup1 mutant with the pollen grains of WT, all the F1 plants exhibited normal fertility and the F2 plants showed a segregation of 249 fertile and 78 sterile plants (3 : 1, χ2 = 0.26, P > 0.05), demonstrating that this male sterile phenotype is controlled by a single recessive gene. To investigate the cytological defects of the nup1 mutant, anthers were analyzed through semi-thin sections based on anther developmental stages as described previously (Zhang et al., 2011). We found that no differences were visible between the nup1 mutant and WT during Stage 7 (S7) to Stage 9 (S9) (Fig. S2a). Notably, the nup1 mutant exhibited obviously shrinking tapetal cells and swollen microspores at S10 and S11, but the swollen microspores rapidly shriveled and crushed at late S11 and S12 (Figs 1f,g, S2b), resulting in no pollen grain production compared with the WT which underwent the regular anther developments.
Transmission electron microscopy (TEM) analysis was performed as described in Methods S1 to observe more detailed structure changes in nup1 anthers. At Stage 8b (S8b), both the nup1 mutant and WT underwent meiosis normally to produce the tetrads, and their tapetal cells were covered by a thin layer of cell wall with no Ubisch body formation (Fig. S3a–f). At Stage 9 (S9), WT tapetal cells started to produce Ubisch bodies, whereas in the nup1 mutant, the Ubisch bodies did not yet emerged on tapetal cells (Fig. S3g–l). At S10, WT tapetal cells were more condensed and formed a large number of characteristic Ubisch bodies on their inner surface, and the pollen walls formed well-organized structures including nexine, bacula and tectum (Figs 1h, S4a). However, interestingly, no Ubisch bodies emerged on the surface of the nup1 tapetum, and the bacula was missing accompanied by narrowed nexine and tectum in the nup1 mutant (Figs 1i, S4b). During S11 to S12, still no Ubisch bodies were observed, and all pollen grains collapsed with tectum and nexine appearing as a much looser two-layer structure in the nup1 mutant (Fig. S4c–n). These results indicated that Ubisch body formation during anther development was completely absent in nup1 mutant anthers. Moreover, scanning electron microscopy (SEM) analysis of S12 anthers showed that the cuticle of WT anther displayed the typical spaghetti-like pattern, whereas the nup1 anther surface lacked the cuticle structure and appeared nearly smooth (Figs 1j,k, S5a,b). In agreement with TEM observations, abundant Ubisch bodies were observed with crystal-like shapes on the locular side of WT anthers, but these structures were completely absent in nup1 anthers (Fig. 1l,m), confirming the loss of Ubisch body in the nup1 mutant. This serious absence of Ubisch body during anther development has not been thoroughly explored in rice. Given these unique characteristics, we thus named this mutant no ubisch body and pollen grain 1 (nup1). Taken together, these results indicate that NUP1 is essentially required for the development of tapetum, anther cuticle, pollen wall and, particularly, the formation of Ubisch bodies.
To test whether the disappearance of the Ubisch body was due to the possible premature degradation of the tapetum in the nup1 mutant, we performed the terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling (TUNEL) assay to detect the timely PCD of the tapetum between WT and the nup1 mutant. In both WT and nup1 anthers, the positive TUNEL signals were clearly visualized at S7–S9 and declined during S10–S11, and there were no obvious differences in TUNEL signals between WT and the nup1 mutant during the anther developmental stages (Fig. S6). Although the cytological analysis showed that the nup1 mutant displayed shrinking tapetal cells at S10 and S11, we did not observe premature degradation of tapetal cells in nup1 mutant anthers compared with WT. These results indicated that loss-of-function of NUP1 did not influence PCD of tapetal cells, but contributed to the loss of Ubisch body formation, which resulted in male sterility in the nup1 mutant.
To clone the NUP1 gene, we generated the F2 population from a cross between the nup1 mutant and indica cv 9311. Using 50 F2 individuals with the nup1 mutant phenotype for bulked segregant analysis by deep sequencing (BSA-seq), the NUP1 locus was limited to a 10.17-Mb region on chromosome 8 (Fig. 1n). By a map-based cloning with designed molecular markers (Table S1), we further narrowed the NUP1 locus to a 121-kb region (Fig. 1o). Whole-genome re-sequencing analysis between WT and the nup1 mutant revealed a single-nucleotide C-to-A substitution in gene coding region within the locus, and this substitution was occurred in the fourth exon of LOC_Os08g20730 (Fig. 1p), resulting in a premature stop codon. Therefore, we identified LOC_Os08g20730 which encodes a class III peroxidase as the candidate gene. To validate this, we developed NUP1 knockout transgenic plants in WT MH86 using the CRISPR/Cas9 approach. Five independent knockout (KO) lines were confirmed to have shifted open reading frames in the first exon (Figs 1p, S7a) and showed smaller and paler anthers with no pollen grains and seed production (Figs 1q,r, S4b–f), identical to the nup1 mutant phenotype. Therefore, we concluded that the NUP1 gene was LOC_Os08g20730.
To determine the expression pattern of NUP1, we performed reverse transcription quantitative polymerase chain reaction and found that NUP1 was preferentially expressed in the anthers during developmental S6 to S12 and lowly expressed in vegetative organs (Fig. 1s). The RNA in situ hybridization of NUP1 in WT showed that NUP1 transcripts were mainly accumulated in tapetal cells and slightly distributed in other cell layers of anther wall and microspores at different developmental stages in anthers (Fig. S8). Furthermore, we generated NUP1-GFP transgenic plants using its native promoter (pNUP1::NUP1-GFP) and observed that NUP1-GFP fusion proteins were substantially localized in all layers of anther wall including tapetum across S7 to S9 (Fig. S9). The rare GFP signal in microspores may be due to a low level of fusion proteins to be observed. Collectively, this expression pattern of NUP1 in anthers confirmed that NUP1 played an essential role in tapetum and microspore development. To determine the subcellular localization of NUP1, we transiently expressed a construct encoding NUP1-GFP fusion protein together with a construct encoding OsH2B-mCherry (histone OsH2B-mCherry fusion, as a nuclear marker) in Nicotiana benthamiana leaves. The fluorescent signals of NUP1-GFP were dominantly localized in the cytoplasm, excluding with OsH2B-mCherry signals (Fig. 1t). The transfection of rice protoplasts with construct encoding NUP1-YFP fusion further confirmed the cytoplasm-localization of NUP1 protein (Fig. S10).
Class III peroxidases are a large family of plant-specific heme-iron–dependent peroxidases that use hydrogen peroxide as the electron acceptor to catalyze a number of oxidative reactions (Shigeto & Tsutsumi, 2016). There are 73 peroxidase genes in Arabidopsis thaliana and 138 genes in rice. These genes are involved in a range of functions, such as lignification, cell elongation and stress defense (Shigeto & Tsutsumi, 2016). Phylogenetic analysis showed that NUP1 was highly conserved among different plant species (Fig. S11). Notably, its closely related homolog in Arabidopsis, PRX40, has been found to function redundantly with PRX9 to maintain tapetum and microspore development (Jacobowitz et al., 2019). Amino acid alignment indicated that the enzyme active sites and heme-ligand binding sites were highly conserved among NUP1, PRX40 and other homologs (Fig. S12). However, even though PRX9 and PRX14, two homologs of rice NUP1 in Arabidopsis have been found to be responsible for Arabidopsis anther development, there are several significant differences in their functions between Arabidopsis and rice: (1) single mutants of either prx9 or prx40 displayed fully fertile, and only prx9 prx40 double-mutant plants were completely sterile in Arabidopsis, whereas single mutant of nup1 exhibits male sterility in rice; (2) tapetal cells of prx9 prx40 double mutants were swollen and invaded into the locular space, while tapetal cells of nup1 in rice were, to some extent, shriveled with absolutely no Ubisch body formation. Therefore, NUP1 and its homologs, PRX9 and PRX14, exhibit some functional diversity in anther development between monocot rice and dicot Arabidopsis.
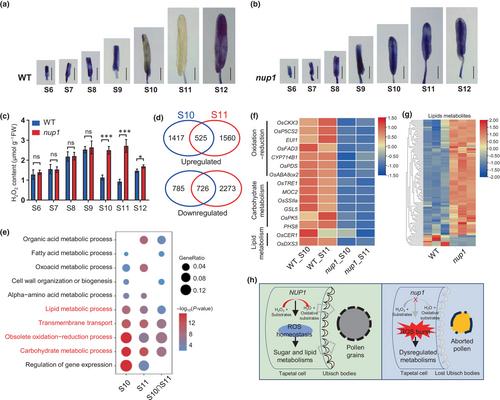
To confirm whether NUP1-mediated ROS scavenging as a peroxidase, we performed nitroblue tetrazolium (NBT) staining to determine the difference in ROS accumulation during anther development between WT and the nup1 mutant. In WT, the ROS signals were clearly detectable during S6 to S9, declined at S10, were almost undetectable at S11, and were re-detectable at S12 (Fig. 2a), which were consistent with previous study (Zhou et al., 2024). During S10 to S11, tapetal cells become degenerated and form Ubisch bodies abundantly, and microspores undergo mitosis, accumulate starch and synthesize the pollen walls using nutrition delivered by Ubisch bodies (Zhang et al., 2011). However, in the nup1 mutant and knockout line KO#1, increased ROS signals were observed at S9, and strong ROS signals were detected at S10 to S11 compared with WT (Figs 2b, S13a,b). Furthermore, we directly measured the contents of H2O2 and malondialdehyde (MDA), which is an indicator of local ROS production and participant in peroxidation of fatty acids, in anthers at various stages. In agreement with the NBT staining, the contents of H2O2 and MDA were comparable in anthers between WT and the nup1 mutant during S6 to S9, whereas they were significantly increased in nup1 anthers compared with WT at S10 and S11 (Figs 2c, S13c), suggesting a ROS burst in nup1 anthers at the later developmental stages. Consistently, a recent study reported that under high temperature excessive ROS in anthers at S10 to S11 in ostms19, tms5, ostms15 and ostms18 is responsible for their high temperature-sensitive male sterility (Zhou et al., 2024). Taking together, these results indicate that the timing of ROS burst in the nup1 mutant corresponds to the defect in nup1 anthers and suggests that NUP1 is involved in ROS scavenging, which is important for Ubisch body formation and microspore development during S10 to S11.
To investigate the impacts of ROS burst on gene expression in nup1 anther, we conducted anther transcriptome profiling (Fig. S14a). Differentially expressed genes (DEGs) analysis (FDR < 0.01, FC > 2) showed that thousands of genes were significantly changed in nup1 anthers compared with WT at S10 and S11 (Fig. S14b,c; Tables S2, S3). We identified 525 upregulated and 726 downregulated DEGs overlapped between anthers at S10 and S11 (Fig. 2d). Gene Ontology (GO) analysis showed that obsolete oxidation–reduction process was the most significantly enriched GO term for DEGs at S10, S11 and the overlapped DEGs at both stages (Fig. 2e), consistent with the ROS burst in the nup1 mutant. Furthermore, the carbohydrate metabolic process, the lipid metabolic process and transmembrane transport were among the top-listed significantly enriched GO terms at both S10 and S11 (Fig. 2e). Indeed, we identified and confirmed that many known genes associated with oxidation–reduction, carbohydrate and lipid metabolisms were significantly up- and downregulated in nup1 anthers compared with WT at both stages (Figs 2d, S14d, S15). Carbohydrate and lipid metabolisms have been widely demonstrated to play critical roles in tapetum development and pollen formation (Wan et al., 2020; Liu et al., 2021). These results indicated that ROS burst in nup1 anthers resulted in defective oxidation–reduction processes and carbohydrate and lipid metabolisms.
To confirm the dysregulated metabolism, we performed mass spectrometry to identify the metabolites in S11 anthers of WT and the nup1 mutant (Fig. S16a). A total of 1963 metabolites including flavonoids, amino acids and derivatives, phenolic acids, lipids, alkaloids, organic acids, lignans and coumarins, and others were identified (Fig. S16b). Differential metabolites analysis showed that 671 compounds were significantly changed in nup1 anthers compared with WT at S11, of which flavonoids (32.34%), lipids (15.80%) and phenolic acids (11.77%) accounted for the top three dysregulated metabolites (Fig. S16c,d; Table S4). In line with the significant GO term related to lipid metabolism, lipid metabolites were predominantly increased in nup1 anthers compared with WT (Fig. 2g). Notably, flavonoids, the antioxidants that mediate direct scavenging of ROS in plants (Agati et al., 2012), were significantly reduced in nup1 anthers (Fig. S16e), This reduction may be attributed to ROS consumption. Moreover, KEGG pathway analysis showed that two lipids, linoleic acid and arachidonic acid metabolism, as well as phenylpropanoid and flavonoid biosynthesis were the significantly enriched terms (Figs S16f, S17). These findings further confirm the dysregulated lipid and oxidation–reduction metabolisms in nup1 anthers.
In summary, we identified the nup1 mutant in rice that exhibited specific male sterility due to the complete absence of Ubisch body formation and pollen grain production. We cloned the causal gene and demonstrated that NUP1, encoding a class III peroxidase predominantly expressed in the anther wall, mediated ROS scavenging. The loss-of-function of NUP1 resulted in ROS burst in anthers, which disrupted the oxidation–reduction process, carbohydrate and lipid metabolisms, and loss of tapetal Ubisch body, thereby resulting in male sterility in rice (Fig. 2h). Our study expands the understanding of the molecular mechanisms by which ROS homeostasis regulates cell metabolism, tapetal Ubisch body formation and male fertility in plants. Recently, the third-generation hybrid rice technology using a recessive nuclear male sterile line has been successfully demonstrated to develop genetic systems for hybrid rice (Chang et al., 2016; Liao et al., 2021). Since the loss of NUP1 resulted in completely absent Ubisch body formation and no pollen grain production without changing female fertility and plant growth, NUP1 is a potentially useful male sterility gene in third-generation hybrid rice breeding in the future.
None declared.
LS, QQ, Y Zhang and HL conceived and designed this research. CS, SY and YC performed most of the experiments. ZX, BZ, MG, Y Zhu and YY planted the materials and performed the gene cloning. CS and YC performed data analysis. CS and SY wrote the manuscript. LS, QQ, Y Zhang, HL and FW provided guidance in writing the manuscript. All authors contributed to editing the manuscript. CS, SY, YC and ZX contributed equally to this work.
期刊介绍:
New Phytologist is an international electronic journal published 24 times a year. It is owned by the New Phytologist Foundation, a non-profit-making charitable organization dedicated to promoting plant science. The journal publishes excellent, novel, rigorous, and timely research and scholarship in plant science and its applications. The articles cover topics in five sections: Physiology & Development, Environment, Interaction, Evolution, and Transformative Plant Biotechnology. These sections encompass intracellular processes, global environmental change, and encourage cross-disciplinary approaches. The journal recognizes the use of techniques from molecular and cell biology, functional genomics, modeling, and system-based approaches in plant science. Abstracting and Indexing Information for New Phytologist includes Academic Search, AgBiotech News & Information, Agroforestry Abstracts, Biochemistry & Biophysics Citation Index, Botanical Pesticides, CAB Abstracts®, Environment Index, Global Health, and Plant Breeding Abstracts, and others.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


