Sara Benedé, Leticia Pérez-Rodríguez, David Menchén-Martínez, Elena Molina, Rosina López-Fandiño
{"title":"气道接触屋尘螨会促进小鼠对蛋白过敏的发展","authors":"Sara Benedé, Leticia Pérez-Rodríguez, David Menchén-Martínez, Elena Molina, Rosina López-Fandiño","doi":"10.1111/cea.14543","DOIUrl":null,"url":null,"abstract":"<p>Cases of adverse reactions to food in young children at their first known exposure have raised intriguing questions about food allergy development, leading to the hypothesis that sensitisation, at least to certain allergens such as peanut, may occur by contact through non-oral pathways like airway inhalation [<span>1</span>]. Interestingly, the biological activity of house dust mite (HDM) allergens stimulates bystander responses to other proteins, which implies that respiratory exposure to food allergens present in domestic dust, such as egg proteins could potentially lead to systemic sensitisation [<span>2</span>]. These findings suggest the possibility that prior sensitisation to egg proteins through the respiratory tract, facilitated by the adjuvant activity of accompanying HDM components, could lead to food allergies when egg is later ingested, in a way similar to that described for peanuts [<span>3, 4</span>]. To test this hypothesis, we used a murine model of sensitisation, without exogenous adjuvants, to investigate the immunostimulant properties of the proteolytically active and inactive forms of HDM in the development of allergy to egg white (EW) when administered either through inhalation or orally. Six-week-old female BALB/c mice received intranasally six doses of HDM, proteolytically inactive HDM (hereafter iHDM), or combinations of EW + HDM and EW + iHDM, followed by eight intragastric gavages with EW, before being intranasally challenged with EW. Materials and methods are provided in the open access repository OSF (https://doi.org/10.17605/OSF.IO/YCNPF).</p><p>Mice that received intranasally EW and its combinations with HDM or iHDM developed EW-specific IgE and IgG1 antibodies, while prior exposure to HDM or iHDM alone did not induce antibodies specific to EW after repeated oral EW administrations over 2 weeks (Figure 1a). Repeated oral administrations of EW did not elicit clinical signs or temperature changes, but, on Day 39, the jejunal concentration of MCP-1 was significantly elevated in mice that had received EW, EW + HDM and EW + iHDM intranasally (Figure 1b), indicating mast cell activation and degranulation in the intestinal mucosa of mice that had produced specific antibodies towards EW. Conversely, intranasal challenge with EW caused anaphylaxis symptoms in the mice that had previously received EW through the airways, particularly in the groups administered EW + HDM and EW + iHDM, which also experienced significant temperature drops, showing that both extracts had adjuvant activity that facilitated airway sensitisation to EW and anaphylaxis following intranasal challenge. Consistently, the concentration of MCP-1 in lung homogenates was significantly elevated in mice exposed to EW + HDM and EW + iHDM (Figure 1b). Mice receiving EW, EW + HDM and EW + iHDM exhibited significantly higher lung levels of IL-6 and IL-4, while the level of TNF-α was significantly higher in the mice exposed to EW + iHDM (Figure 1b). Analyses in the lungs of the expression of genes encoding tight junction proteins revealed that <i>Cldn3</i> (claudin 3) and <i>Tjp1</i> (zonula occludens 1) were upregulated in the mouse groups that received HDM, either alone or in combination with EW (Figure 1c). However, the expression of genes encoding epithelial alarmins, such as <i>Il33</i> (IL-33), <i>Il25</i> (IL-25) and <i>Tslp</i> (TSLP), as well as <i>Il6</i> (IL-6), <i>Il17</i> (IL-17) and <i>Gata3</i> (GATA3) remained similar in all the experimental groups (not shown). Evaluation of dendritic cell (DC) genes involved in the stimulation of Th2 responses in lung tissues revealed that <i>Irf4</i> (IRF4, interferon regulatory factor 4) was differentially enhanced in the mice that had received EW + HDM (Figure 1c). After stimulation with concanavalin A, spleen cells of mice exposed to EW + HDM and EW + iHDM produced IL-5 and IL-13, and those exposed to EW + HDM also significantly released IL-4 to the culture medium (Figure 1d), showing that intranasal exposure to EW in conjunction with HDM or iHDM promoted the development of Th2 responses.</p><p>The adjuvant activity of HDM has been attributed to the degradation of tight junction proteins in airway epithelial cells by HDM proteases [<span>5, 6</span>]. In murine lungs, this induces the production of innate cytokines, which, when combined with co-exposed peanut, promote activation and migration of DCs to lung-draining lymph nodes, development of Th2 responses and production specific IgE, ultimately triggering systemic peanut allergy [<span>3</span>]. However, these barrier effects are at least partially reversible, as <i>de novo</i> protein synthesis helps to maintain junctional integrity [<span>7</span>]. This explains the upregulation of the expression of genes encoding tight junction proteins in our study, that would act as a compensatory mechanism to improve epithelial function. Similarly, the fact that we did not detect changes in the gene expression of alarmins in lung tissues probably reflects the time elapsed since the last intranasal administrations of the combinations of EW with either HDM or iHDM. In addition to proteolytic activity, the activation of innate immunity via TLR4 and TLR2 signalling by compounds present in HDM is known to initiate Th2 polarisation and HDM allergy [<span>8, 9</span>]. Notably, EW + iHDM significantly increased the level of TNF-α in lung homogenates, that arises from activation of inflammatory pathways downstream of the common TLR adaptor MyD88. In agreement with our results, Smeekens et al. [<span>4</span>] found that proteolytic activity plays a minor role in the adjuvant activity of household dust, concluding that TLR ligands that activate MyD88 signalling are the crucial components responsible for its immunomodulatory properties that promote sensitisation to simultaneously inhaled peanut in mice.</p><p>In conclusion, HDM extracts, whether proteolytically active or inactive, enhanced the development of a Th2-driven immune response to EW through the airways, leading to allergic manifestations. The presence of active proteases in HDM extracts slightly increased their adjuvant effect, suggesting that protease activity played a modest role in this context.</p><p>Study concept and design, analysis and interpretation of data and drafting of the manuscript: Sara Benedé and Rosina López-Fandiño. Active conduct of the experiments: Sara Benedé, David Menchén-Martínez and Leticia Pérez-Rodríguez. Review and revision of the manuscript: Sara Benedé, Elena Molina and Rosina López-Fandiño. Obtained funding: Elena Molina and Rosina López-Fandiño.</p><p>All protocols followed the European Legislation (directive 2010/63/EU) and were approved by the Comunidad de Madrid (Ref PROEX 286.8/20).</p><p>The authors declare no conflicts of interest.</p>","PeriodicalId":10207,"journal":{"name":"Clinical and Experimental Allergy","volume":"54 10","pages":"777-780"},"PeriodicalIF":6.3000,"publicationDate":"2024-07-15","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1111/cea.14543","citationCount":"0","resultStr":"{\"title\":\"Airway Exposure to House Dust Mite Promotes the Development of Allergy to Egg White in Mice\",\"authors\":\"Sara Benedé, Leticia Pérez-Rodríguez, David Menchén-Martínez, Elena Molina, Rosina López-Fandiño\",\"doi\":\"10.1111/cea.14543\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p>Cases of adverse reactions to food in young children at their first known exposure have raised intriguing questions about food allergy development, leading to the hypothesis that sensitisation, at least to certain allergens such as peanut, may occur by contact through non-oral pathways like airway inhalation [<span>1</span>]. Interestingly, the biological activity of house dust mite (HDM) allergens stimulates bystander responses to other proteins, which implies that respiratory exposure to food allergens present in domestic dust, such as egg proteins could potentially lead to systemic sensitisation [<span>2</span>]. These findings suggest the possibility that prior sensitisation to egg proteins through the respiratory tract, facilitated by the adjuvant activity of accompanying HDM components, could lead to food allergies when egg is later ingested, in a way similar to that described for peanuts [<span>3, 4</span>]. To test this hypothesis, we used a murine model of sensitisation, without exogenous adjuvants, to investigate the immunostimulant properties of the proteolytically active and inactive forms of HDM in the development of allergy to egg white (EW) when administered either through inhalation or orally. Six-week-old female BALB/c mice received intranasally six doses of HDM, proteolytically inactive HDM (hereafter iHDM), or combinations of EW + HDM and EW + iHDM, followed by eight intragastric gavages with EW, before being intranasally challenged with EW. Materials and methods are provided in the open access repository OSF (https://doi.org/10.17605/OSF.IO/YCNPF).</p><p>Mice that received intranasally EW and its combinations with HDM or iHDM developed EW-specific IgE and IgG1 antibodies, while prior exposure to HDM or iHDM alone did not induce antibodies specific to EW after repeated oral EW administrations over 2 weeks (Figure 1a). Repeated oral administrations of EW did not elicit clinical signs or temperature changes, but, on Day 39, the jejunal concentration of MCP-1 was significantly elevated in mice that had received EW, EW + HDM and EW + iHDM intranasally (Figure 1b), indicating mast cell activation and degranulation in the intestinal mucosa of mice that had produced specific antibodies towards EW. Conversely, intranasal challenge with EW caused anaphylaxis symptoms in the mice that had previously received EW through the airways, particularly in the groups administered EW + HDM and EW + iHDM, which also experienced significant temperature drops, showing that both extracts had adjuvant activity that facilitated airway sensitisation to EW and anaphylaxis following intranasal challenge. Consistently, the concentration of MCP-1 in lung homogenates was significantly elevated in mice exposed to EW + HDM and EW + iHDM (Figure 1b). Mice receiving EW, EW + HDM and EW + iHDM exhibited significantly higher lung levels of IL-6 and IL-4, while the level of TNF-α was significantly higher in the mice exposed to EW + iHDM (Figure 1b). Analyses in the lungs of the expression of genes encoding tight junction proteins revealed that <i>Cldn3</i> (claudin 3) and <i>Tjp1</i> (zonula occludens 1) were upregulated in the mouse groups that received HDM, either alone or in combination with EW (Figure 1c). However, the expression of genes encoding epithelial alarmins, such as <i>Il33</i> (IL-33), <i>Il25</i> (IL-25) and <i>Tslp</i> (TSLP), as well as <i>Il6</i> (IL-6), <i>Il17</i> (IL-17) and <i>Gata3</i> (GATA3) remained similar in all the experimental groups (not shown). Evaluation of dendritic cell (DC) genes involved in the stimulation of Th2 responses in lung tissues revealed that <i>Irf4</i> (IRF4, interferon regulatory factor 4) was differentially enhanced in the mice that had received EW + HDM (Figure 1c). After stimulation with concanavalin A, spleen cells of mice exposed to EW + HDM and EW + iHDM produced IL-5 and IL-13, and those exposed to EW + HDM also significantly released IL-4 to the culture medium (Figure 1d), showing that intranasal exposure to EW in conjunction with HDM or iHDM promoted the development of Th2 responses.</p><p>The adjuvant activity of HDM has been attributed to the degradation of tight junction proteins in airway epithelial cells by HDM proteases [<span>5, 6</span>]. In murine lungs, this induces the production of innate cytokines, which, when combined with co-exposed peanut, promote activation and migration of DCs to lung-draining lymph nodes, development of Th2 responses and production specific IgE, ultimately triggering systemic peanut allergy [<span>3</span>]. However, these barrier effects are at least partially reversible, as <i>de novo</i> protein synthesis helps to maintain junctional integrity [<span>7</span>]. This explains the upregulation of the expression of genes encoding tight junction proteins in our study, that would act as a compensatory mechanism to improve epithelial function. Similarly, the fact that we did not detect changes in the gene expression of alarmins in lung tissues probably reflects the time elapsed since the last intranasal administrations of the combinations of EW with either HDM or iHDM. In addition to proteolytic activity, the activation of innate immunity via TLR4 and TLR2 signalling by compounds present in HDM is known to initiate Th2 polarisation and HDM allergy [<span>8, 9</span>]. Notably, EW + iHDM significantly increased the level of TNF-α in lung homogenates, that arises from activation of inflammatory pathways downstream of the common TLR adaptor MyD88. In agreement with our results, Smeekens et al. [<span>4</span>] found that proteolytic activity plays a minor role in the adjuvant activity of household dust, concluding that TLR ligands that activate MyD88 signalling are the crucial components responsible for its immunomodulatory properties that promote sensitisation to simultaneously inhaled peanut in mice.</p><p>In conclusion, HDM extracts, whether proteolytically active or inactive, enhanced the development of a Th2-driven immune response to EW through the airways, leading to allergic manifestations. The presence of active proteases in HDM extracts slightly increased their adjuvant effect, suggesting that protease activity played a modest role in this context.</p><p>Study concept and design, analysis and interpretation of data and drafting of the manuscript: Sara Benedé and Rosina López-Fandiño. Active conduct of the experiments: Sara Benedé, David Menchén-Martínez and Leticia Pérez-Rodríguez. Review and revision of the manuscript: Sara Benedé, Elena Molina and Rosina López-Fandiño. Obtained funding: Elena Molina and Rosina López-Fandiño.</p><p>All protocols followed the European Legislation (directive 2010/63/EU) and were approved by the Comunidad de Madrid (Ref PROEX 286.8/20).</p><p>The authors declare no conflicts of interest.</p>\",\"PeriodicalId\":10207,\"journal\":{\"name\":\"Clinical and Experimental Allergy\",\"volume\":\"54 10\",\"pages\":\"777-780\"},\"PeriodicalIF\":6.3000,\"publicationDate\":\"2024-07-15\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://onlinelibrary.wiley.com/doi/epdf/10.1111/cea.14543\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"Clinical and Experimental Allergy\",\"FirstCategoryId\":\"3\",\"ListUrlMain\":\"https://onlinelibrary.wiley.com/doi/10.1111/cea.14543\",\"RegionNum\":2,\"RegionCategory\":\"医学\",\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"Q1\",\"JCRName\":\"ALLERGY\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"Clinical and Experimental Allergy","FirstCategoryId":"3","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1111/cea.14543","RegionNum":2,"RegionCategory":"医学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"ALLERGY","Score":null,"Total":0}
引用次数: 0
摘要
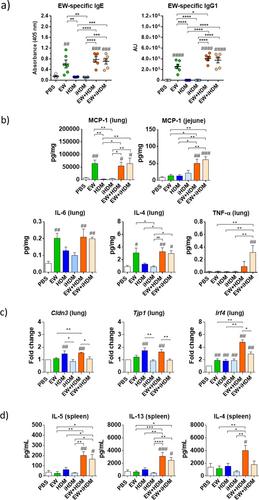
幼儿在首次接触食物时出现不良反应的病例引发了有关食物过敏发展的有趣问题,从而提出了这样的假设:至少对某些过敏原(如花生)的过敏可能是通过非口途径(如气道吸入)接触引起的[1]。有趣的是,屋尘螨(HDM)过敏原的生物活性会刺激旁观者对其他蛋白质产生反应,这意味着通过呼吸道接触家庭灰尘中的食物过敏原(如鸡蛋蛋白)有可能导致全身过敏[2]。这些研究结果表明,通过呼吸道对鸡蛋蛋白产生过敏之前,由于伴随的人类发展指数(HDM)成分的佐剂作用,可能会导致后来摄入鸡蛋时产生食物过敏,其方式与花生过敏的方式类似[3, 4]。为了验证这一假设,我们使用了一种不含外源性佐剂的小鼠致敏模型,以研究蛋白水解活性和非活性形式的 HDM 在通过吸入或口服给药导致对蛋白(EW)过敏的过程中的免疫刺激特性。六周大的雌性 BALB/c 小鼠在鼻腔内接受六次 HDM、蛋白水解非活性 HDM(以下简称 iHDM)或 EW + HDM 和 EW + iHDM 的组合剂量,然后在胃内灌胃八次 EW,最后接受 EW 的鼻腔挑战。小鼠鼻内注射EW及其与HDM或iHDM的组合后会产生EW特异性IgE和IgG1抗体,而之前单独接触HDM或iHDM的小鼠在2周内重复口服EW后不会诱发EW特异性抗体(图1a)。重复口服 EW 不会引起临床症状或体温变化,但在第 39 天,鼻内注射 EW、EW + HDM 和 EW + iHDM 的小鼠空肠 MCP-1 浓度显著升高(图 1b),这表明产生 EW 特异性抗体的小鼠肠粘膜肥大细胞活化和脱颗粒。相反,鼻内挑战 EW 会导致之前通过气道接受 EW 的小鼠出现过敏性休克症状,特别是在注射 EW + HDM 和 EW + iHDM 的组别中,体温也会显著下降,这表明这两种提取物都具有佐剂活性,可促进气道对 EW 的过敏和鼻内挑战后的过敏性休克。同样,暴露于 EW + HDM 和 EW + iHDM 的小鼠肺匀浆中的 MCP-1 浓度显著升高(图 1b)。接受 EW、EW + HDM 和 EW + iHDM 的小鼠肺部 IL-6 和 IL-4 水平明显升高,而 TNF-α 水平在接触 EW + iHDM 的小鼠中明显升高(图 1b)。对肺部紧密连接蛋白编码基因的表达进行分析后发现,在单独或与 EW 一起接受 HDM 的小鼠组中,Cldn3(claudin 3)和 Tjp1(zonula occludens 1)的表达上调(图 1c)。然而,在所有实验组中,编码上皮alarmins的基因,如Il33(IL-33)、Il25(IL-25)和Tslp(TSLP),以及Il6(IL-6)、Il17(IL-17)和Gata3(GATA3)的表达仍然相似(未显示)。对参与刺激肺组织 Th2 反应的树突状细胞(DC)基因的评估显示,在接受 EW + HDM 的小鼠中,Irf4(IRF4,干扰素调节因子 4)不同程度地增强(图 1c)。暴露于EW+HDM和EW+iHDM的小鼠脾脏细胞在受到Concanavalin A刺激后产生了IL-5和IL-13,而暴露于EW+HDM的小鼠脾脏细胞也向培养基显著释放了IL-4(图1d),这表明鼻内暴露于EW与HDM或iHDM可促进Th2反应的发展。HDM的佐剂活性是由于HDM蛋白酶降解了气道上皮细胞中的紧密连接蛋白[5,6]。在小鼠肺中,这会诱导先天性细胞因子的产生,当这些细胞因子与共同接触的花生结合时,会促进 DCs 的活化和向肺排泄淋巴结的迁移、Th2 反应的发展以及特异性 IgE 的产生,最终引发全身性花生过敏[3]。然而,这些屏障效应至少部分是可逆的,因为新蛋白质的合成有助于维持连接的完整性[7]。这就解释了为什么在我们的研究中编码紧密连接蛋白的基因表达上调,这将作为一种改善上皮功能的补偿机制。同样,我们没有在肺组织中检测到allarmins基因表达的变化,这可能反映了自上一次鼻内给药EW与HDM或iHDM的组合后已经过去了一段时间。
Airway Exposure to House Dust Mite Promotes the Development of Allergy to Egg White in Mice
Cases of adverse reactions to food in young children at their first known exposure have raised intriguing questions about food allergy development, leading to the hypothesis that sensitisation, at least to certain allergens such as peanut, may occur by contact through non-oral pathways like airway inhalation [1]. Interestingly, the biological activity of house dust mite (HDM) allergens stimulates bystander responses to other proteins, which implies that respiratory exposure to food allergens present in domestic dust, such as egg proteins could potentially lead to systemic sensitisation [2]. These findings suggest the possibility that prior sensitisation to egg proteins through the respiratory tract, facilitated by the adjuvant activity of accompanying HDM components, could lead to food allergies when egg is later ingested, in a way similar to that described for peanuts [3, 4]. To test this hypothesis, we used a murine model of sensitisation, without exogenous adjuvants, to investigate the immunostimulant properties of the proteolytically active and inactive forms of HDM in the development of allergy to egg white (EW) when administered either through inhalation or orally. Six-week-old female BALB/c mice received intranasally six doses of HDM, proteolytically inactive HDM (hereafter iHDM), or combinations of EW + HDM and EW + iHDM, followed by eight intragastric gavages with EW, before being intranasally challenged with EW. Materials and methods are provided in the open access repository OSF (https://doi.org/10.17605/OSF.IO/YCNPF).
Mice that received intranasally EW and its combinations with HDM or iHDM developed EW-specific IgE and IgG1 antibodies, while prior exposure to HDM or iHDM alone did not induce antibodies specific to EW after repeated oral EW administrations over 2 weeks (Figure 1a). Repeated oral administrations of EW did not elicit clinical signs or temperature changes, but, on Day 39, the jejunal concentration of MCP-1 was significantly elevated in mice that had received EW, EW + HDM and EW + iHDM intranasally (Figure 1b), indicating mast cell activation and degranulation in the intestinal mucosa of mice that had produced specific antibodies towards EW. Conversely, intranasal challenge with EW caused anaphylaxis symptoms in the mice that had previously received EW through the airways, particularly in the groups administered EW + HDM and EW + iHDM, which also experienced significant temperature drops, showing that both extracts had adjuvant activity that facilitated airway sensitisation to EW and anaphylaxis following intranasal challenge. Consistently, the concentration of MCP-1 in lung homogenates was significantly elevated in mice exposed to EW + HDM and EW + iHDM (Figure 1b). Mice receiving EW, EW + HDM and EW + iHDM exhibited significantly higher lung levels of IL-6 and IL-4, while the level of TNF-α was significantly higher in the mice exposed to EW + iHDM (Figure 1b). Analyses in the lungs of the expression of genes encoding tight junction proteins revealed that Cldn3 (claudin 3) and Tjp1 (zonula occludens 1) were upregulated in the mouse groups that received HDM, either alone or in combination with EW (Figure 1c). However, the expression of genes encoding epithelial alarmins, such as Il33 (IL-33), Il25 (IL-25) and Tslp (TSLP), as well as Il6 (IL-6), Il17 (IL-17) and Gata3 (GATA3) remained similar in all the experimental groups (not shown). Evaluation of dendritic cell (DC) genes involved in the stimulation of Th2 responses in lung tissues revealed that Irf4 (IRF4, interferon regulatory factor 4) was differentially enhanced in the mice that had received EW + HDM (Figure 1c). After stimulation with concanavalin A, spleen cells of mice exposed to EW + HDM and EW + iHDM produced IL-5 and IL-13, and those exposed to EW + HDM also significantly released IL-4 to the culture medium (Figure 1d), showing that intranasal exposure to EW in conjunction with HDM or iHDM promoted the development of Th2 responses.
The adjuvant activity of HDM has been attributed to the degradation of tight junction proteins in airway epithelial cells by HDM proteases [5, 6]. In murine lungs, this induces the production of innate cytokines, which, when combined with co-exposed peanut, promote activation and migration of DCs to lung-draining lymph nodes, development of Th2 responses and production specific IgE, ultimately triggering systemic peanut allergy [3]. However, these barrier effects are at least partially reversible, as de novo protein synthesis helps to maintain junctional integrity [7]. This explains the upregulation of the expression of genes encoding tight junction proteins in our study, that would act as a compensatory mechanism to improve epithelial function. Similarly, the fact that we did not detect changes in the gene expression of alarmins in lung tissues probably reflects the time elapsed since the last intranasal administrations of the combinations of EW with either HDM or iHDM. In addition to proteolytic activity, the activation of innate immunity via TLR4 and TLR2 signalling by compounds present in HDM is known to initiate Th2 polarisation and HDM allergy [8, 9]. Notably, EW + iHDM significantly increased the level of TNF-α in lung homogenates, that arises from activation of inflammatory pathways downstream of the common TLR adaptor MyD88. In agreement with our results, Smeekens et al. [4] found that proteolytic activity plays a minor role in the adjuvant activity of household dust, concluding that TLR ligands that activate MyD88 signalling are the crucial components responsible for its immunomodulatory properties that promote sensitisation to simultaneously inhaled peanut in mice.
In conclusion, HDM extracts, whether proteolytically active or inactive, enhanced the development of a Th2-driven immune response to EW through the airways, leading to allergic manifestations. The presence of active proteases in HDM extracts slightly increased their adjuvant effect, suggesting that protease activity played a modest role in this context.
Study concept and design, analysis and interpretation of data and drafting of the manuscript: Sara Benedé and Rosina López-Fandiño. Active conduct of the experiments: Sara Benedé, David Menchén-Martínez and Leticia Pérez-Rodríguez. Review and revision of the manuscript: Sara Benedé, Elena Molina and Rosina López-Fandiño. Obtained funding: Elena Molina and Rosina López-Fandiño.
All protocols followed the European Legislation (directive 2010/63/EU) and were approved by the Comunidad de Madrid (Ref PROEX 286.8/20).
期刊介绍:
Clinical & Experimental Allergy strikes an excellent balance between clinical and scientific articles and carries regular reviews and editorials written by leading authorities in their field.
In response to the increasing number of quality submissions, since 1996 the journals size has increased by over 30%. Clinical & Experimental Allergy is essential reading for allergy practitioners and research scientists with an interest in allergic diseases and mechanisms. Truly international in appeal, Clinical & Experimental Allergy publishes clinical and experimental observations in disease in all fields of medicine in which allergic hypersensitivity plays a part.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


