Fatma İşbilir, Cansel Güzin Özgüden Akkoç, İhsan İşbilir, Özgür Özöner, Barış Can Güzel, İlker Arıcan, Şükrü Hakan Atalgın, Mehmet Can, Soner Bekmezci
{"title":"火鸡(Meleagris gallopavo)泌尿生殖腺的形态、宏观和微观研究。","authors":"Fatma İşbilir, Cansel Güzin Özgüden Akkoç, İhsan İşbilir, Özgür Özöner, Barış Can Güzel, İlker Arıcan, Şükrü Hakan Atalgın, Mehmet Can, Soner Bekmezci","doi":"10.1002/jemt.24642","DOIUrl":null,"url":null,"abstract":"<p>Birds lack sebaceous and sweat glands, whereas mammals have abundant eccrine, apocrine, and sebaceous glands. The glandulae uropygiale (preen gland) of birds is a unique structure that may resemble the sebaceous glands of mammals (Jacob & Ziswiler, <span>1982</span>; King & McLelland, <span>1985</span>; Reynolds, <span>2013</span>).</p><p>A study of 126 species, 49 families in total, reports that the uropygial gland is present in phylogenetically distant birds (Tinamidae-Hirundinidae) or, conversely, is absent in phylogenetically close taxa (some <i>Psittacidae</i> and <i>Columbidae</i>), according to the phylogenetic classification of birds (Morony et al., <span>1975</span>), the developmental trend of the gland is independent of the lineage of birds (Montalti et al., <span>2000</span>).</p><p>Studies have shown that the uropygial gland is more developed in waterfowl than in land birds (Farner et al., <span>1982</span>). The gland is topographically located on the last caudal vertebra (Johnston, <span>1988</span>) and in the region between the fourth caudal vertebra and the pygostyle (Lucas & Stettenheim, <span>1972</span>; Yılmaz et al., <span>2018</span>). It has also been reported to be located on the last sacral and first caudal vertebrae in quail (Atalgın & Kürtül, <span>2008</span>) and neighboring the caudal muscles in quail and turkeys (Farner et al., <span>1982</span>; Özcan et al., <span>2004</span>; Rajathi, <span>2019</span>).</p><p>The gland usually consists of two lobes and a duct system (Carril et al., <span>2019</span>; Çınar & Kelek, <span>2014</span>; Rajathi, <span>2019</span>). The lobes are separated from each other by the interlobular septum from the inside (Carril et al., <span>2019</span>; Haydar, <span>2005</span>; Yılmaz et al., <span>2018</span>). The duct system transfers the produced oily secretion to a typical nipple-like papilla located on the tail (King & McLelland, <span>1985</span>). It has a tuft of feather around the papilla (Lucas & Stettenheim, <span>1972</span>; Schumacher, <span>1919</span>). The shape and size of the gland differ between species (Jacob & Ziswiler, <span>1982</span>).</p><p>The secretion of the gland consists of oil-synthesizing enzymes, cell debris, and volatile substances (Bhattacacharyya & Ghosh, <span>1971</span>; Burger et al., <span>2004</span>; Campagna et al., <span>2012</span>). Secretion content varies according to sex, age, diet, and sexual period (Sandilands et al., <span>2004</span>; Serbest, <span>2004</span>; Zık & Erdost, <span>2002</span>).</p><p>Although the function of the gland is controversial, its secretion has been reported to have effects such as feather care (Chiale et al., <span>2021</span>; Reynolds, <span>2013</span>), indirect cell regeneration, cell growth, and differentiation (Liu, Qi, et al., <span>2022</span>), antibacterial (Chiale et al., <span>2020</span>; Galván et al., <span>2008</span>; Shawkey et al., <span>2003</span>), pheromone-like in sexual behavior (Liu et al., <span>2022a</span>; Zhang et al., <span>2010</span>), protection against predators, or parasites (Caspers et al., <span>2021</span>).</p><p>Histologically, the gland is surrounded externally by a dense connective tissue capsule (Carril et al., <span>2019</span>; Chiale et al., <span>2016</span>). Gland epithelium consists of different cellular layers in the form of a germinative layer, intermediate layer, secretory layer, and degenerative layer (Carril et al., <span>2019</span>; Jacob & Zeman, <span>1972</span>). Some studies have reported that the gland is a compound tubulo-alveolar holocrine, ectodermal invagination (Jacob & Ziswiler, <span>1982</span>; Sandilands et al., <span>2004</span>; Wagner & Boord, <span>1975</span>). In addition, it has been reported that the gland in quail, roosters, Aseel breed roosters and chickens, and ducks has a simple tubular structure with holocrine-type secretion (Çınar & Kelek, <span>2014</span>; Harem et al., <span>2005</span>; Suzuki & Kusuhara, <span>1996</span>; Yılmaz et al., <span>2018</span>; Zık & Erdost, <span>2002</span>).</p><p>The turkey (<i>Meleagris gallopavo</i>) is a bird in the order <i>Galliformes</i>, family Phasanidae. It has been recognized as a species with several subspecies of its wild ancestor spread over a wide area from southern Canada to southern Mexico. It was reported to have been domesticated before Christopher Columbus discovered the Americas (Crawford, <span>1992</span>; Thornton, <span>2016</span>). The structure of the uropygial gland in some bird species in the order <i>Galliformes</i> has been studied macroscopically and microscopically (Çınar & Kelek, <span>2014</span>; Demırcıoglu et al., <span>2019</span>; Rajathi, <span>2019</span>). Apart from routine triple and Hematoxylin & Eosin (H&E) stains, histochemical stains are available (Chiale et al., <span>2016</span>, <span>2019</span>; Çınar & Kelek, <span>2014</span>). In addition, scanning electron microscopy (SEM) studies are quite limited. In the literature, there is a study on macroscopic and some morphometric features of the uropygial gland in turkeys (Rajathi, <span>2019</span>). However, no macroscopic, morphometric, or microscopic examination of this gland about sex differences was found. This study is the first study to investigate macroscopically, morphometrically, and microscopically the uropygial gland in turkeys depending on sex. Microscopic examination was performed using light and SEM.</p><p><i>Glandulae uropygialis</i> of a total of 13 turkeys (7 females and 6 males) were used as material in the study. The glands were obtained from turkeys that did not show any clinical findings and died from reasons unrelated to the study. The tissues were delivered fresh to the laboratory. The turkeys were brought to the laboratory in autumn and no evidence of a reproductive cycle was observed.</p><p>In our study, the presence of the uropygial gland was observed in all turkeys. The gland was topographically located on the last caudal vertebrae. It was covered with a thin skin (Figure 2a). The gland was observed to be embedded in a layer of adipose tissue (Figure 2b). It was determined that the gland consisted of two lobes and a papilla system (Figure 2c,d). The lobes had a symmetrical appearance from an external view. The dorsal and ventral surfaces showed a distinct convexity. It was observed that it resembled the heart in shape and the ends of the lobes merged caudally. There was a short papilla at the caudal end of the gland. Numerous holes (porus glandulae uropygialis) were observed on the papilla, which allowed the secretion to drain out. In addition, many feather structures (Figure 2c,d) were observed in these holes. The gland was divided into two lobes by a <i>septum interlobare</i> (Figure 3). In addition, the presence of ducts named ductus glandulae uropygialis was determined under a stereomicroscope (Figure 3). Inside the lobes, a brown secretion material with a dense consistency attracted attention. The general characterization of glandula uropygialis in turkeys was presented in Table 1.</p><p>In male and female turkeys, uropygial gland weights were measured as 2.50 ± 0.16 g and 3.04 ± 0.21 g, respectively. As a result of the statistical evaluation, glandula uropygialis lobe length, papilla width, and papilla height (<i>p</i> < .05) parameters were significantly different between the sexes. It was determined that glandula uropygialis lobe width, papilla width, and papilla height parameters were larger in male turkeys than in females. At the same time, as a result of the index calculations, it was determined that there was no difference between the sexes in terms RGWI, LGUI, and GULI, while PUI value was higher in male turkeys and showed a statistical difference between the sexes (<i>p</i> < .05). Descriptive statistical values and <i>p</i> values of measurement parameters and index values are presented in Tables 2 and 3, respectively.</p><p>The first histological finding was that the glandula uropygialis was embedded in a dense adipose tissue (Figure 4). In each of the histological staining, both female and male turkeys' uropygial glands histological structures were shown the same structures. The gland was a bi-lobed structure surrounded by a thick fibrous capsule. There were blood and lymph vessels, nerve fibers, adipocytes, and smooth muscle bands in the capsule. The capsule was divided the gland ventrally down between the lobes to form the interlobular septum where blood and lymph vessels, nerve fibers, and adipocytes were located (Figures 5 and 6). The gland structure was determined to be tubular. It was seen to have a simple branched tubular structure in some areas, but mostly a simple tubular gland structure. Each lobe of the organ consisted of several secretory tubules. All tubules were separated from each other by a thin connective tissue. The secretory tubules were divided into central and peripheral tubules. Although the lumens of the central tubules were wide, no distinct lumen structure was observed in the peripheral tubules (Figure 7). Secretory products were observed in some of the central tubule's lumen (Figures 7 and 8). The central and peripheral secretory tubules were composed of four distinct layers differing in shape and cell arrangement basal cell layer, intermediate cell layer, secretory cell layer, and degenerative cell layer (Figure 8). The intermediate cell layer consisted of 2–3 layers of polyhedral cells with eosinophilic cytoplasm and centrally located nuclei. Different-sized, vacuolated, polyhedral cells, and containing secretory granules, were observed to form the secretory cell layer. The degenerated cell layer adjacent to the tubule lumen was composed of cells with pyknotic nuclei and pale cytoplasm (Figure 8). In the parts of the interlobular septum close to the papilla, dense amounts of adipose tissue, nerve fibers, and smooth muscle structures were observed. In our study, lymphoid tissue cell communities were located both between the tubular glands (Figure 7a) and in the connective tissue forming the interlobular septum (Figure 6b). The excrete ducts opened into two big collector ducts (ductus glandulae uropygialis) caudal to the gland (Figure 9a,b). PAS (+) reaction was observed in the basal membrane and secretory material (Figure 8b). The fact that the reaction was not glycogen-induced was determined by the observation of PAS positivity as a result of staining with the PAS-diastase technique (Figure 10a). As a result of Orcein staining, elastic fibers were found in the capsule surrounding the uropygial gland and in the intertubular and interlobular septum (Figure 10b).</p><p>SEM revealed the presence of a thick connective tissue capsule outside the uropygial gland (Figure 11b,c). Glandular lobes consisted of lobules variable in size and shape (Figure 11). The lobules consisted of different shaped follicles (sphere-like) protruding from the surface of the gland and marked by a thick band of connective tissue on the outside (Figure 12a,b). It was observed that these glandular follicle structures were gathered together in different shapes or mostly disorganized. It was also observed that these follicle structures were connected to each other by connective tissue threads (interfollicular septa) (Figure 12a). The duct opening of each lobe was separated from the gland surface by a shallow groove containing a mass of lipid secretion radiating from the gland surface towards the ductal wall. This opening was observed to be oval in shape, of varying thickness and with a folded edge (Figure 12c). Multiple holes (porus glandulae uropygialis<i>)</i> were observed caudal to the papilla (Figure 12d).</p><p>Although the glandula uropygialis is present in almost all bird species during the embryonic period, it may be vestigialised in adults of some orders, families, genera, and species (Johnston, <span>1988</span>; Salibian & Montalti, <span>2009</span>). In our study, uropygial gland was present in all turkeys examined.</p><p>It has been reported that the gland is located in the region between the fourth caudal vertebra and the pygostyle in different bird species (Lucas & Stettenheim, <span>1972</span>; Yılmaz et al., <span>2018</span>), at the base of the tail, on the pygostyle muscles. (Johnston, <span>1988</span>; Sawad, <span>2006</span>) Between the caudal side of the lumbosacral bone and the first coccygeal vertebra (Kozlu et al., <span>2011</span>) and generally on the free caudal vertebrae (Moreno-Rueda, <span>2016</span>; Yılmaz & Yılmaz, <span>2019</span>). Our study determined that the uropygial gland was located on the last caudal vertebra in the literature.</p><p>Similar to Rajathi (<span>2019</span>) and Lucas & Stettenheim, <span>1972</span> (fowl and quail), it was determined as a two-lobed gland. The uropygial gland in this study had two lobes, right and left, and a common papilla with ducts draining secretion from these glands. The lobes were orientated forwards and outwards. The lobes of the uropygial gland were pear-shaped, as in turkeys and chickens (Farner et al., <span>1982</span>; Rajathi, <span>2019</span>).</p><p>The common papilla of the gland was thick and elongated as reported in domestic chickens (Lucas & Stettenheim, <span>1972</span>; Rajathi, <span>2019</span>). The papillae were surrounded by skin covered with small feathers extending dorsocaudally, as in chickens (Lucas & Stettenheim, <span>1972</span>).</p><p>In domestic chickens, it has been reported that the ducts open outwards with a breast-like papilla located in the midline, caudal, and dorsal (Gezici, <span>2002</span>). The papilla structure was reported to be conical in <i>Psittaciformes</i> species and cylindrical in Monk parrots (Carril et al., <span>2019</span>). In this study, the papilla structure of the gland was determined to be single, thick, and long, similar to domestic chickens (Lucas & Stettenheim, <span>1972</span>) and turkeys (Rajathi, <span>2019</span>). In our study, it was determined that the papilla had two separate main ducts coming from each lobe. Similar results have been found in poultry and Japanese quail (Farner et al., <span>1982</span>; Mobini & Ziaii, <span>2011</span>; Rajathi, <span>2019</span>). In addition, similar to studies in chickens and roosters (Yılmaz & Yılmaz, <span>2019</span>), it was found that numerous feather structures surrounded the papilla. Numerous holes (porus glandulae uropygialis) were observed at the caudal end of the papilla structure. These holes allow the secretion of secretion from the two uropygial gland ducts.</p><p>Age, season, and sex have been found to have a significant effect on the weight of the uropygial gland (Chen et al., <span>2015</span>; Elder, <span>1954</span>). In our study, the weight of uropygial glands in male and female turkeys was found to be 2.50 ± 0.16 g and 3.04 ± 0.21 g, respectively. The same value was reported as 0.95 ± 0.15 and 0.91 ± 0.26 g in Aseel roosters and chicks, respectively (Yılmaz et al., <span>2018</span>). Demırcıoglu et al. (<span>2019</span>) reported the relative gland weight as 0.11 ± 0.01 g in quails. The average value of RGWI in Aseel breed hens and roosters was found to be 0.0423 (Yılmaz et al., <span>2018</span>) and 0.094 ± 0.041 in <i>Gallus gallus</i> (Montalti et al., <span>2000</span>). Rajathi (<span>2019</span>) determined this index value as 0.01 ± 0.01 in turkeys, while 0.03 ± 0.001 was found in our study. There was no statistical difference between male and female animals in terms of RGWI value.</p><p>Rajathi (<span>2019</span>) reported right and left lobe length parameters in turkeys as 9.7 ± 0.2 mm and 9.6 ± 0.2 mm, respectively. In a study conducted in quails, right GUL was determined as 10.31 mm in females and 10.39 mm in males (Demırcıoglu et al., <span>2019</span>). In our study, GUL parameter was found to be higher than the values determined by Rajathi. In addition, while there was a statistical difference between male and female animals, the value was higher in male animals. Similarly, PUH and PUW parameters had a statistically significant difference between male and female animals in our study. PUH value was higher in quail (Demırcıoglu et al., <span>2019</span>), Aseel breed roosters and hens (Yılmaz et al., <span>2018</span>) and turkeys (Rajathi, <span>2019</span>). In our study, it was determined that these parameters were greater in male turkeys than females.</p><p>Histologic examination revealed that the glandula uropygialis was surrounded by a capsule consisting of a thick connective tissue layer on the outside, similar to studies in quail, partridge, and chickens. There are two lobes and a papilla system under the capsule (Demırcıoglu et al., <span>2019</span>; Önal et al., <span>2013</span>; Yılmaz & Yılmaz, <span>2020</span>). The capsule structure contained blood and lymph vessels, nerve fibers, adipocytes, and smooth muscle bands. Kozlu et al. (<span>2011</span>) and Chiale et al. (<span>2016</span>) reported that the excretory ducts opening the glands were located in the center of the lobe. In our study, similar to the report of Yılmaz and Yılmaz (<span>2020</span>), ducts opened into two main excretory ducts (ductus glandulae uropygialis) caudal to the gland. It is consistent with the literature that the secretory tubules of both lobes are divided into central and peripheral tubules and located around a central lumen (Balkaya et al., <span>2016</span>; Kozlu et al., <span>2011</span>; Önal et al., <span>2013</span>; Stangier et al., <span>2023</span>). Contrary to the studies reporting that the gland has a compound tubulo-alveolar holocrine character (Jacob & Ziswiler, <span>1982</span>; Sandilands et al., <span>2004</span>; Wagner & Boord, <span>1975</span>), in our study, it was determined that it has a simple tubular structure with holocrine secretion as reported in many studies (Çınar & Kelek, <span>2014</span>; Harem et al., <span>2005</span>; Suzuki & Kusuhara, <span>1996</span>; Yılmaz et al., <span>2018</span>; Zık & Erdost, <span>2002</span>).</p><p>Similar to studies in quails and guinea fowl, in our study, central and peripheral secretory tubules were found to consist of four different cell layers differing in shape and cell arrangement from periphery to center: basal cell layer, intermediate cell layer, secretory cell layer, and degenerative cell layer (Ushakumary et al., <span>2011</span>; Zık & Erdost, <span>2002</span>). Although the lumens of the central tubules were wide, no distinct lumen structure was observed in the peripheral tubules.</p><p>The presence of smooth muscle cells, blood, and lymph vessels in the structure of the gland, especially in the <i>septum interlobare</i> and intertubular septum, has been reported (Yılmaz et al., <span>2018</span>). Similarly, smooth muscle cells were found especially in <i>trabeculae</i> in magpies (Balkaya et al., <span>2016</span>). However, some studies have reported that the glandula uropygialis of poultry species such as geese (Hou, <span>1928</span>), grouse (Sawad, <span>2006</span>), and starlings (Sadoon, <span>2011</span>) are completely devoid of smooth muscle cells. In our study, smooth muscle cells were observed in the regions of the interlobular septum close to the papilla. It was determined that the intertubular septum was composed of connective tissue and did not contain smooth muscle. Blood and lymph vessels, nerve fibers, and adipocytes were also observed in the interlobular septum.</p><p>In a study in Pekin ducks, it was reported that small basal cells with flat nuclei located on the basement membrane, large cells filled with lipid droplets towards the lumen of the tubules and cells with pyknotic nuclei and large lipid droplets close to the lumen were found in the gland (Carpenter & Goodridge, <span>1988</span>). In roosters, it was determined as flat nuclei with acidophilic cytoplasm located on the basement membrane (Zık & Erdost, <span>2002</span>). In our study, the basal cell layer consisted of cells with darkly stained nuclei and basophilic cytoplasm. Yılmaz et al. (<span>2018</span>) reported that intermediate cells have acidophilic cytoplasm and basophilic nuclei, secretory cells have many large and small, white-colored fat vacuoles in the lumen, while degenerative cells have larger fat vacuoles in the cytoplasm. In our study, the intermediate cell layer consisted of 2–3 rows of centrally located polyhedral cells with eosinophilic cytoplasm. The secretory cell layer was composed of polyhedral cells of different sizes with vacuoles and secretory granules. The degenerated cell layer adjacent to the tubule lumen consisted of cells with pyknotic nuclei and pale cytoplasm. Acidophilic cells were also detected in H&E staining. The high number of acidophilic cells indicates that there is a large amount of fat synthesis in the gland (Yılmaz et al., <span>2018</span>). Herbst corpuscles, which were first identified in wild ducks (Harem et al., <span>2005</span>), were also determined in laughing pigeons (Madkour et al., <span>2023</span>). Similar to the study in mallard ducks (Yılmaz & Yılmaz, <span>2019</span>), Herbst corpuscles structure was not detected in our study.</p><p>In a study on the rooster uropygial gland, the apical half of the tubules, which were found to be rich in glycogen by the PAS staining method, was named as glycogen region and it was reported that this region gave a strong PAS (+) reaction (Zık & Erdost, <span>2002</span>). In the female and male partridge uropygial gland, no PAS (+) reaction was observed in the sebaceous region, while in females, it was determined that neutral mucosubstance reacted with moderate intensity in the peripheral parts of some tubules belonging to the sebaceous region. In addition, a strong PAS reaction was detected in females and a moderate to strong PAS reaction was detected in males in the inter-tubular area of the fat region (Önal et al., <span>2013</span>). In Chilean Flamingo and Monk parakeets, a positive reaction in the uropygial glands with PAS and Alcian Blue (pH 2.5) revealed the presence of glycoconjugates containing carboxyl groups and sulfated esters. These compounds have been associated with protective functions (protection and maintenance of feathers) in digestive tract organs in fish (Díaz et al., <span>2008</span>; Yashpal et al., <span>2014</span>) and uropygial glands in birds (Carril et al., <span>2019</span>; Chiale et al., <span>2021</span>). In our study, positive staining with PAS was observed especially in the basement membrane and secretory material. After PAS diastase staining, similar to Chiale et al. (<span>2016</span>), it was determined that the staining was not glycogen-induced. As a result of Orcein staining, elastic fibers were observed in the capsule and intertubular septum, which is consistent with the literature (Carril et al., <span>2019</span>; Chiale et al., <span>2016</span>, <span>2020</span>, <span>2021</span>).</p><p>Studies have reported the presence of lymphoid cell communities between tubular glands (Harem et al., <span>2005</span>; Sawad, <span>2006</span>; Shafiian & Mobini, <span>2014</span>). It has also been reported that lymphoid cell communities are located in the connective tissue and papillae at the junction of the glands and excretory duct (Chiale et al., <span>2016</span>; Yılmaz & Yılmaz, <span>2020</span>). In our study, it was determined that lymphoid tissue cell communities were located both between tubular glands and in connective tissue. Also contains abundant dense connective tissue, smooth muscle fibers, and lymphoid tissue.</p><p>Madkour et al., <span>2023</span> reported that the glandular lobe consisted of elliptical, triangular, round glandular lobules, and glandular follicle structures came together in different shapes as a result of SEM analysis. In our study, glandular lobules were spherical and elliptical, and glandular follicle structures were mostly scattered. While the duct opening was reported to be oval shaped, with curved edges, and symmetrical thickness Madkour et al. (<span>2023</span>), in our study it was found to be oval shaped, of variable thickness, and with curved edges. There was no difference in the histologic and electron microscopic structure of the glandula uropygialis in male and female turkeys used in the study.</p><p>Although a study was conducted by Rajathi in 2019 to determine morphometric features in turkeys, this is the first study to perform light and electron microscopic identification of the glandula uropygialis in turkeys. The macroanatomical, morphometric, and microscopic findings of the glandula uropygialis in the turkeys used in the study were found to be very similar to those of most avian species, but different from those of aquatic species. The differences may be due to species and habitat differences.</p><p><b>Fatma İşbilir:</b> Data curation; investigation; validation; formal analysis; supervision; conceptualization; methodology; resources; writing – original draft; software; visualization; project administration; writing – review and editing. <b>Cansel Güzin Özgüden Akkoç:</b> Methodology; formal analysis; supervision; writing – original draft; writing – review and editing; visualization. <b>İhsan İşbilir:</b> Methodology; visualization; writing – original draft; writing – review and editing; formal analysis; supervision. <b>Özgür Özöner:</b> Methodology; supervision; visualization; writing – review and editing; writing – original draft; formal analysis. <b>Barış Can Güzel:</b> Methodology; formal analysis; supervision; writing – original draft; writing – review and editing. <b>İlker ARICAN:</b> Methodology; formal analysis; supervision; writing – original draft; writing – review and editing. <b>Şükrü Hakan Atalgın:</b> Visualization; writing – original draft; writing – review and editing; conceptualization; methodology. <b>Mehmet Can:</b> Conceptualization; methodology; visualization; writing – review and editing; writing – original draft. <b>Soner Bekmezci:</b> Conceptualization; methodology; visualization; writing – review and editing; writing – original draft.</p>","PeriodicalId":2,"journal":{"name":"ACS Applied Bio Materials","volume":null,"pages":null},"PeriodicalIF":4.6000,"publicationDate":"2024-06-28","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1002/jemt.24642","citationCount":"0","resultStr":"{\"title\":\"Morphometric, macroscopic and microscopic investigation of glandulae uropygiale in turkeys (Meleagris gallopavo)\",\"authors\":\"Fatma İşbilir, Cansel Güzin Özgüden Akkoç, İhsan İşbilir, Özgür Özöner, Barış Can Güzel, İlker Arıcan, Şükrü Hakan Atalgın, Mehmet Can, Soner Bekmezci\",\"doi\":\"10.1002/jemt.24642\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p>Birds lack sebaceous and sweat glands, whereas mammals have abundant eccrine, apocrine, and sebaceous glands. The glandulae uropygiale (preen gland) of birds is a unique structure that may resemble the sebaceous glands of mammals (Jacob & Ziswiler, <span>1982</span>; King & McLelland, <span>1985</span>; Reynolds, <span>2013</span>).</p><p>A study of 126 species, 49 families in total, reports that the uropygial gland is present in phylogenetically distant birds (Tinamidae-Hirundinidae) or, conversely, is absent in phylogenetically close taxa (some <i>Psittacidae</i> and <i>Columbidae</i>), according to the phylogenetic classification of birds (Morony et al., <span>1975</span>), the developmental trend of the gland is independent of the lineage of birds (Montalti et al., <span>2000</span>).</p><p>Studies have shown that the uropygial gland is more developed in waterfowl than in land birds (Farner et al., <span>1982</span>). The gland is topographically located on the last caudal vertebra (Johnston, <span>1988</span>) and in the region between the fourth caudal vertebra and the pygostyle (Lucas & Stettenheim, <span>1972</span>; Yılmaz et al., <span>2018</span>). It has also been reported to be located on the last sacral and first caudal vertebrae in quail (Atalgın & Kürtül, <span>2008</span>) and neighboring the caudal muscles in quail and turkeys (Farner et al., <span>1982</span>; Özcan et al., <span>2004</span>; Rajathi, <span>2019</span>).</p><p>The gland usually consists of two lobes and a duct system (Carril et al., <span>2019</span>; Çınar & Kelek, <span>2014</span>; Rajathi, <span>2019</span>). The lobes are separated from each other by the interlobular septum from the inside (Carril et al., <span>2019</span>; Haydar, <span>2005</span>; Yılmaz et al., <span>2018</span>). The duct system transfers the produced oily secretion to a typical nipple-like papilla located on the tail (King & McLelland, <span>1985</span>). It has a tuft of feather around the papilla (Lucas & Stettenheim, <span>1972</span>; Schumacher, <span>1919</span>). The shape and size of the gland differ between species (Jacob & Ziswiler, <span>1982</span>).</p><p>The secretion of the gland consists of oil-synthesizing enzymes, cell debris, and volatile substances (Bhattacacharyya & Ghosh, <span>1971</span>; Burger et al., <span>2004</span>; Campagna et al., <span>2012</span>). Secretion content varies according to sex, age, diet, and sexual period (Sandilands et al., <span>2004</span>; Serbest, <span>2004</span>; Zık & Erdost, <span>2002</span>).</p><p>Although the function of the gland is controversial, its secretion has been reported to have effects such as feather care (Chiale et al., <span>2021</span>; Reynolds, <span>2013</span>), indirect cell regeneration, cell growth, and differentiation (Liu, Qi, et al., <span>2022</span>), antibacterial (Chiale et al., <span>2020</span>; Galván et al., <span>2008</span>; Shawkey et al., <span>2003</span>), pheromone-like in sexual behavior (Liu et al., <span>2022a</span>; Zhang et al., <span>2010</span>), protection against predators, or parasites (Caspers et al., <span>2021</span>).</p><p>Histologically, the gland is surrounded externally by a dense connective tissue capsule (Carril et al., <span>2019</span>; Chiale et al., <span>2016</span>). Gland epithelium consists of different cellular layers in the form of a germinative layer, intermediate layer, secretory layer, and degenerative layer (Carril et al., <span>2019</span>; Jacob & Zeman, <span>1972</span>). Some studies have reported that the gland is a compound tubulo-alveolar holocrine, ectodermal invagination (Jacob & Ziswiler, <span>1982</span>; Sandilands et al., <span>2004</span>; Wagner & Boord, <span>1975</span>). In addition, it has been reported that the gland in quail, roosters, Aseel breed roosters and chickens, and ducks has a simple tubular structure with holocrine-type secretion (Çınar & Kelek, <span>2014</span>; Harem et al., <span>2005</span>; Suzuki & Kusuhara, <span>1996</span>; Yılmaz et al., <span>2018</span>; Zık & Erdost, <span>2002</span>).</p><p>The turkey (<i>Meleagris gallopavo</i>) is a bird in the order <i>Galliformes</i>, family Phasanidae. It has been recognized as a species with several subspecies of its wild ancestor spread over a wide area from southern Canada to southern Mexico. It was reported to have been domesticated before Christopher Columbus discovered the Americas (Crawford, <span>1992</span>; Thornton, <span>2016</span>). The structure of the uropygial gland in some bird species in the order <i>Galliformes</i> has been studied macroscopically and microscopically (Çınar & Kelek, <span>2014</span>; Demırcıoglu et al., <span>2019</span>; Rajathi, <span>2019</span>). Apart from routine triple and Hematoxylin & Eosin (H&E) stains, histochemical stains are available (Chiale et al., <span>2016</span>, <span>2019</span>; Çınar & Kelek, <span>2014</span>). In addition, scanning electron microscopy (SEM) studies are quite limited. In the literature, there is a study on macroscopic and some morphometric features of the uropygial gland in turkeys (Rajathi, <span>2019</span>). However, no macroscopic, morphometric, or microscopic examination of this gland about sex differences was found. This study is the first study to investigate macroscopically, morphometrically, and microscopically the uropygial gland in turkeys depending on sex. Microscopic examination was performed using light and SEM.</p><p><i>Glandulae uropygialis</i> of a total of 13 turkeys (7 females and 6 males) were used as material in the study. The glands were obtained from turkeys that did not show any clinical findings and died from reasons unrelated to the study. The tissues were delivered fresh to the laboratory. The turkeys were brought to the laboratory in autumn and no evidence of a reproductive cycle was observed.</p><p>In our study, the presence of the uropygial gland was observed in all turkeys. The gland was topographically located on the last caudal vertebrae. It was covered with a thin skin (Figure 2a). The gland was observed to be embedded in a layer of adipose tissue (Figure 2b). It was determined that the gland consisted of two lobes and a papilla system (Figure 2c,d). The lobes had a symmetrical appearance from an external view. The dorsal and ventral surfaces showed a distinct convexity. It was observed that it resembled the heart in shape and the ends of the lobes merged caudally. There was a short papilla at the caudal end of the gland. Numerous holes (porus glandulae uropygialis) were observed on the papilla, which allowed the secretion to drain out. In addition, many feather structures (Figure 2c,d) were observed in these holes. The gland was divided into two lobes by a <i>septum interlobare</i> (Figure 3). In addition, the presence of ducts named ductus glandulae uropygialis was determined under a stereomicroscope (Figure 3). Inside the lobes, a brown secretion material with a dense consistency attracted attention. The general characterization of glandula uropygialis in turkeys was presented in Table 1.</p><p>In male and female turkeys, uropygial gland weights were measured as 2.50 ± 0.16 g and 3.04 ± 0.21 g, respectively. As a result of the statistical evaluation, glandula uropygialis lobe length, papilla width, and papilla height (<i>p</i> < .05) parameters were significantly different between the sexes. It was determined that glandula uropygialis lobe width, papilla width, and papilla height parameters were larger in male turkeys than in females. At the same time, as a result of the index calculations, it was determined that there was no difference between the sexes in terms RGWI, LGUI, and GULI, while PUI value was higher in male turkeys and showed a statistical difference between the sexes (<i>p</i> < .05). Descriptive statistical values and <i>p</i> values of measurement parameters and index values are presented in Tables 2 and 3, respectively.</p><p>The first histological finding was that the glandula uropygialis was embedded in a dense adipose tissue (Figure 4). In each of the histological staining, both female and male turkeys' uropygial glands histological structures were shown the same structures. The gland was a bi-lobed structure surrounded by a thick fibrous capsule. There were blood and lymph vessels, nerve fibers, adipocytes, and smooth muscle bands in the capsule. The capsule was divided the gland ventrally down between the lobes to form the interlobular septum where blood and lymph vessels, nerve fibers, and adipocytes were located (Figures 5 and 6). The gland structure was determined to be tubular. It was seen to have a simple branched tubular structure in some areas, but mostly a simple tubular gland structure. Each lobe of the organ consisted of several secretory tubules. All tubules were separated from each other by a thin connective tissue. The secretory tubules were divided into central and peripheral tubules. Although the lumens of the central tubules were wide, no distinct lumen structure was observed in the peripheral tubules (Figure 7). Secretory products were observed in some of the central tubule's lumen (Figures 7 and 8). The central and peripheral secretory tubules were composed of four distinct layers differing in shape and cell arrangement basal cell layer, intermediate cell layer, secretory cell layer, and degenerative cell layer (Figure 8). The intermediate cell layer consisted of 2–3 layers of polyhedral cells with eosinophilic cytoplasm and centrally located nuclei. Different-sized, vacuolated, polyhedral cells, and containing secretory granules, were observed to form the secretory cell layer. The degenerated cell layer adjacent to the tubule lumen was composed of cells with pyknotic nuclei and pale cytoplasm (Figure 8). In the parts of the interlobular septum close to the papilla, dense amounts of adipose tissue, nerve fibers, and smooth muscle structures were observed. In our study, lymphoid tissue cell communities were located both between the tubular glands (Figure 7a) and in the connective tissue forming the interlobular septum (Figure 6b). The excrete ducts opened into two big collector ducts (ductus glandulae uropygialis) caudal to the gland (Figure 9a,b). PAS (+) reaction was observed in the basal membrane and secretory material (Figure 8b). The fact that the reaction was not glycogen-induced was determined by the observation of PAS positivity as a result of staining with the PAS-diastase technique (Figure 10a). As a result of Orcein staining, elastic fibers were found in the capsule surrounding the uropygial gland and in the intertubular and interlobular septum (Figure 10b).</p><p>SEM revealed the presence of a thick connective tissue capsule outside the uropygial gland (Figure 11b,c). Glandular lobes consisted of lobules variable in size and shape (Figure 11). The lobules consisted of different shaped follicles (sphere-like) protruding from the surface of the gland and marked by a thick band of connective tissue on the outside (Figure 12a,b). It was observed that these glandular follicle structures were gathered together in different shapes or mostly disorganized. It was also observed that these follicle structures were connected to each other by connective tissue threads (interfollicular septa) (Figure 12a). The duct opening of each lobe was separated from the gland surface by a shallow groove containing a mass of lipid secretion radiating from the gland surface towards the ductal wall. This opening was observed to be oval in shape, of varying thickness and with a folded edge (Figure 12c). Multiple holes (porus glandulae uropygialis<i>)</i> were observed caudal to the papilla (Figure 12d).</p><p>Although the glandula uropygialis is present in almost all bird species during the embryonic period, it may be vestigialised in adults of some orders, families, genera, and species (Johnston, <span>1988</span>; Salibian & Montalti, <span>2009</span>). In our study, uropygial gland was present in all turkeys examined.</p><p>It has been reported that the gland is located in the region between the fourth caudal vertebra and the pygostyle in different bird species (Lucas & Stettenheim, <span>1972</span>; Yılmaz et al., <span>2018</span>), at the base of the tail, on the pygostyle muscles. (Johnston, <span>1988</span>; Sawad, <span>2006</span>) Between the caudal side of the lumbosacral bone and the first coccygeal vertebra (Kozlu et al., <span>2011</span>) and generally on the free caudal vertebrae (Moreno-Rueda, <span>2016</span>; Yılmaz & Yılmaz, <span>2019</span>). Our study determined that the uropygial gland was located on the last caudal vertebra in the literature.</p><p>Similar to Rajathi (<span>2019</span>) and Lucas & Stettenheim, <span>1972</span> (fowl and quail), it was determined as a two-lobed gland. The uropygial gland in this study had two lobes, right and left, and a common papilla with ducts draining secretion from these glands. The lobes were orientated forwards and outwards. The lobes of the uropygial gland were pear-shaped, as in turkeys and chickens (Farner et al., <span>1982</span>; Rajathi, <span>2019</span>).</p><p>The common papilla of the gland was thick and elongated as reported in domestic chickens (Lucas & Stettenheim, <span>1972</span>; Rajathi, <span>2019</span>). The papillae were surrounded by skin covered with small feathers extending dorsocaudally, as in chickens (Lucas & Stettenheim, <span>1972</span>).</p><p>In domestic chickens, it has been reported that the ducts open outwards with a breast-like papilla located in the midline, caudal, and dorsal (Gezici, <span>2002</span>). The papilla structure was reported to be conical in <i>Psittaciformes</i> species and cylindrical in Monk parrots (Carril et al., <span>2019</span>). In this study, the papilla structure of the gland was determined to be single, thick, and long, similar to domestic chickens (Lucas & Stettenheim, <span>1972</span>) and turkeys (Rajathi, <span>2019</span>). In our study, it was determined that the papilla had two separate main ducts coming from each lobe. Similar results have been found in poultry and Japanese quail (Farner et al., <span>1982</span>; Mobini & Ziaii, <span>2011</span>; Rajathi, <span>2019</span>). In addition, similar to studies in chickens and roosters (Yılmaz & Yılmaz, <span>2019</span>), it was found that numerous feather structures surrounded the papilla. Numerous holes (porus glandulae uropygialis) were observed at the caudal end of the papilla structure. These holes allow the secretion of secretion from the two uropygial gland ducts.</p><p>Age, season, and sex have been found to have a significant effect on the weight of the uropygial gland (Chen et al., <span>2015</span>; Elder, <span>1954</span>). In our study, the weight of uropygial glands in male and female turkeys was found to be 2.50 ± 0.16 g and 3.04 ± 0.21 g, respectively. The same value was reported as 0.95 ± 0.15 and 0.91 ± 0.26 g in Aseel roosters and chicks, respectively (Yılmaz et al., <span>2018</span>). Demırcıoglu et al. (<span>2019</span>) reported the relative gland weight as 0.11 ± 0.01 g in quails. The average value of RGWI in Aseel breed hens and roosters was found to be 0.0423 (Yılmaz et al., <span>2018</span>) and 0.094 ± 0.041 in <i>Gallus gallus</i> (Montalti et al., <span>2000</span>). Rajathi (<span>2019</span>) determined this index value as 0.01 ± 0.01 in turkeys, while 0.03 ± 0.001 was found in our study. There was no statistical difference between male and female animals in terms of RGWI value.</p><p>Rajathi (<span>2019</span>) reported right and left lobe length parameters in turkeys as 9.7 ± 0.2 mm and 9.6 ± 0.2 mm, respectively. In a study conducted in quails, right GUL was determined as 10.31 mm in females and 10.39 mm in males (Demırcıoglu et al., <span>2019</span>). In our study, GUL parameter was found to be higher than the values determined by Rajathi. In addition, while there was a statistical difference between male and female animals, the value was higher in male animals. Similarly, PUH and PUW parameters had a statistically significant difference between male and female animals in our study. PUH value was higher in quail (Demırcıoglu et al., <span>2019</span>), Aseel breed roosters and hens (Yılmaz et al., <span>2018</span>) and turkeys (Rajathi, <span>2019</span>). In our study, it was determined that these parameters were greater in male turkeys than females.</p><p>Histologic examination revealed that the glandula uropygialis was surrounded by a capsule consisting of a thick connective tissue layer on the outside, similar to studies in quail, partridge, and chickens. There are two lobes and a papilla system under the capsule (Demırcıoglu et al., <span>2019</span>; Önal et al., <span>2013</span>; Yılmaz & Yılmaz, <span>2020</span>). The capsule structure contained blood and lymph vessels, nerve fibers, adipocytes, and smooth muscle bands. Kozlu et al. (<span>2011</span>) and Chiale et al. (<span>2016</span>) reported that the excretory ducts opening the glands were located in the center of the lobe. In our study, similar to the report of Yılmaz and Yılmaz (<span>2020</span>), ducts opened into two main excretory ducts (ductus glandulae uropygialis) caudal to the gland. It is consistent with the literature that the secretory tubules of both lobes are divided into central and peripheral tubules and located around a central lumen (Balkaya et al., <span>2016</span>; Kozlu et al., <span>2011</span>; Önal et al., <span>2013</span>; Stangier et al., <span>2023</span>). Contrary to the studies reporting that the gland has a compound tubulo-alveolar holocrine character (Jacob & Ziswiler, <span>1982</span>; Sandilands et al., <span>2004</span>; Wagner & Boord, <span>1975</span>), in our study, it was determined that it has a simple tubular structure with holocrine secretion as reported in many studies (Çınar & Kelek, <span>2014</span>; Harem et al., <span>2005</span>; Suzuki & Kusuhara, <span>1996</span>; Yılmaz et al., <span>2018</span>; Zık & Erdost, <span>2002</span>).</p><p>Similar to studies in quails and guinea fowl, in our study, central and peripheral secretory tubules were found to consist of four different cell layers differing in shape and cell arrangement from periphery to center: basal cell layer, intermediate cell layer, secretory cell layer, and degenerative cell layer (Ushakumary et al., <span>2011</span>; Zık & Erdost, <span>2002</span>). Although the lumens of the central tubules were wide, no distinct lumen structure was observed in the peripheral tubules.</p><p>The presence of smooth muscle cells, blood, and lymph vessels in the structure of the gland, especially in the <i>septum interlobare</i> and intertubular septum, has been reported (Yılmaz et al., <span>2018</span>). Similarly, smooth muscle cells were found especially in <i>trabeculae</i> in magpies (Balkaya et al., <span>2016</span>). However, some studies have reported that the glandula uropygialis of poultry species such as geese (Hou, <span>1928</span>), grouse (Sawad, <span>2006</span>), and starlings (Sadoon, <span>2011</span>) are completely devoid of smooth muscle cells. In our study, smooth muscle cells were observed in the regions of the interlobular septum close to the papilla. It was determined that the intertubular septum was composed of connective tissue and did not contain smooth muscle. Blood and lymph vessels, nerve fibers, and adipocytes were also observed in the interlobular septum.</p><p>In a study in Pekin ducks, it was reported that small basal cells with flat nuclei located on the basement membrane, large cells filled with lipid droplets towards the lumen of the tubules and cells with pyknotic nuclei and large lipid droplets close to the lumen were found in the gland (Carpenter & Goodridge, <span>1988</span>). In roosters, it was determined as flat nuclei with acidophilic cytoplasm located on the basement membrane (Zık & Erdost, <span>2002</span>). In our study, the basal cell layer consisted of cells with darkly stained nuclei and basophilic cytoplasm. Yılmaz et al. (<span>2018</span>) reported that intermediate cells have acidophilic cytoplasm and basophilic nuclei, secretory cells have many large and small, white-colored fat vacuoles in the lumen, while degenerative cells have larger fat vacuoles in the cytoplasm. In our study, the intermediate cell layer consisted of 2–3 rows of centrally located polyhedral cells with eosinophilic cytoplasm. The secretory cell layer was composed of polyhedral cells of different sizes with vacuoles and secretory granules. The degenerated cell layer adjacent to the tubule lumen consisted of cells with pyknotic nuclei and pale cytoplasm. Acidophilic cells were also detected in H&E staining. The high number of acidophilic cells indicates that there is a large amount of fat synthesis in the gland (Yılmaz et al., <span>2018</span>). Herbst corpuscles, which were first identified in wild ducks (Harem et al., <span>2005</span>), were also determined in laughing pigeons (Madkour et al., <span>2023</span>). Similar to the study in mallard ducks (Yılmaz & Yılmaz, <span>2019</span>), Herbst corpuscles structure was not detected in our study.</p><p>In a study on the rooster uropygial gland, the apical half of the tubules, which were found to be rich in glycogen by the PAS staining method, was named as glycogen region and it was reported that this region gave a strong PAS (+) reaction (Zık & Erdost, <span>2002</span>). In the female and male partridge uropygial gland, no PAS (+) reaction was observed in the sebaceous region, while in females, it was determined that neutral mucosubstance reacted with moderate intensity in the peripheral parts of some tubules belonging to the sebaceous region. In addition, a strong PAS reaction was detected in females and a moderate to strong PAS reaction was detected in males in the inter-tubular area of the fat region (Önal et al., <span>2013</span>). In Chilean Flamingo and Monk parakeets, a positive reaction in the uropygial glands with PAS and Alcian Blue (pH 2.5) revealed the presence of glycoconjugates containing carboxyl groups and sulfated esters. These compounds have been associated with protective functions (protection and maintenance of feathers) in digestive tract organs in fish (Díaz et al., <span>2008</span>; Yashpal et al., <span>2014</span>) and uropygial glands in birds (Carril et al., <span>2019</span>; Chiale et al., <span>2021</span>). In our study, positive staining with PAS was observed especially in the basement membrane and secretory material. After PAS diastase staining, similar to Chiale et al. (<span>2016</span>), it was determined that the staining was not glycogen-induced. As a result of Orcein staining, elastic fibers were observed in the capsule and intertubular septum, which is consistent with the literature (Carril et al., <span>2019</span>; Chiale et al., <span>2016</span>, <span>2020</span>, <span>2021</span>).</p><p>Studies have reported the presence of lymphoid cell communities between tubular glands (Harem et al., <span>2005</span>; Sawad, <span>2006</span>; Shafiian & Mobini, <span>2014</span>). It has also been reported that lymphoid cell communities are located in the connective tissue and papillae at the junction of the glands and excretory duct (Chiale et al., <span>2016</span>; Yılmaz & Yılmaz, <span>2020</span>). In our study, it was determined that lymphoid tissue cell communities were located both between tubular glands and in connective tissue. Also contains abundant dense connective tissue, smooth muscle fibers, and lymphoid tissue.</p><p>Madkour et al., <span>2023</span> reported that the glandular lobe consisted of elliptical, triangular, round glandular lobules, and glandular follicle structures came together in different shapes as a result of SEM analysis. In our study, glandular lobules were spherical and elliptical, and glandular follicle structures were mostly scattered. While the duct opening was reported to be oval shaped, with curved edges, and symmetrical thickness Madkour et al. (<span>2023</span>), in our study it was found to be oval shaped, of variable thickness, and with curved edges. There was no difference in the histologic and electron microscopic structure of the glandula uropygialis in male and female turkeys used in the study.</p><p>Although a study was conducted by Rajathi in 2019 to determine morphometric features in turkeys, this is the first study to perform light and electron microscopic identification of the glandula uropygialis in turkeys. The macroanatomical, morphometric, and microscopic findings of the glandula uropygialis in the turkeys used in the study were found to be very similar to those of most avian species, but different from those of aquatic species. The differences may be due to species and habitat differences.</p><p><b>Fatma İşbilir:</b> Data curation; investigation; validation; formal analysis; supervision; conceptualization; methodology; resources; writing – original draft; software; visualization; project administration; writing – review and editing. <b>Cansel Güzin Özgüden Akkoç:</b> Methodology; formal analysis; supervision; writing – original draft; writing – review and editing; visualization. <b>İhsan İşbilir:</b> Methodology; visualization; writing – original draft; writing – review and editing; formal analysis; supervision. <b>Özgür Özöner:</b> Methodology; supervision; visualization; writing – review and editing; writing – original draft; formal analysis. <b>Barış Can Güzel:</b> Methodology; formal analysis; supervision; writing – original draft; writing – review and editing. <b>İlker ARICAN:</b> Methodology; formal analysis; supervision; writing – original draft; writing – review and editing. <b>Şükrü Hakan Atalgın:</b> Visualization; writing – original draft; writing – review and editing; conceptualization; methodology. <b>Mehmet Can:</b> Conceptualization; methodology; visualization; writing – review and editing; writing – original draft. <b>Soner Bekmezci:</b> Conceptualization; methodology; visualization; writing – review and editing; writing – original draft.</p>\",\"PeriodicalId\":2,\"journal\":{\"name\":\"ACS Applied Bio Materials\",\"volume\":null,\"pages\":null},\"PeriodicalIF\":4.6000,\"publicationDate\":\"2024-06-28\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://onlinelibrary.wiley.com/doi/epdf/10.1002/jemt.24642\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"ACS Applied Bio Materials\",\"FirstCategoryId\":\"5\",\"ListUrlMain\":\"https://onlinelibrary.wiley.com/doi/10.1002/jemt.24642\",\"RegionNum\":0,\"RegionCategory\":null,\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"Q2\",\"JCRName\":\"MATERIALS SCIENCE, BIOMATERIALS\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"ACS Applied Bio Materials","FirstCategoryId":"5","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1002/jemt.24642","RegionNum":0,"RegionCategory":null,"ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q2","JCRName":"MATERIALS SCIENCE, BIOMATERIALS","Score":null,"Total":0}
Morphometric, macroscopic and microscopic investigation of glandulae uropygiale in turkeys (Meleagris gallopavo)
Birds lack sebaceous and sweat glands, whereas mammals have abundant eccrine, apocrine, and sebaceous glands. The glandulae uropygiale (preen gland) of birds is a unique structure that may resemble the sebaceous glands of mammals (Jacob & Ziswiler, 1982; King & McLelland, 1985; Reynolds, 2013).
A study of 126 species, 49 families in total, reports that the uropygial gland is present in phylogenetically distant birds (Tinamidae-Hirundinidae) or, conversely, is absent in phylogenetically close taxa (some Psittacidae and Columbidae), according to the phylogenetic classification of birds (Morony et al., 1975), the developmental trend of the gland is independent of the lineage of birds (Montalti et al., 2000).
Studies have shown that the uropygial gland is more developed in waterfowl than in land birds (Farner et al., 1982). The gland is topographically located on the last caudal vertebra (Johnston, 1988) and in the region between the fourth caudal vertebra and the pygostyle (Lucas & Stettenheim, 1972; Yılmaz et al., 2018). It has also been reported to be located on the last sacral and first caudal vertebrae in quail (Atalgın & Kürtül, 2008) and neighboring the caudal muscles in quail and turkeys (Farner et al., 1982; Özcan et al., 2004; Rajathi, 2019).
The gland usually consists of two lobes and a duct system (Carril et al., 2019; Çınar & Kelek, 2014; Rajathi, 2019). The lobes are separated from each other by the interlobular septum from the inside (Carril et al., 2019; Haydar, 2005; Yılmaz et al., 2018). The duct system transfers the produced oily secretion to a typical nipple-like papilla located on the tail (King & McLelland, 1985). It has a tuft of feather around the papilla (Lucas & Stettenheim, 1972; Schumacher, 1919). The shape and size of the gland differ between species (Jacob & Ziswiler, 1982).
The secretion of the gland consists of oil-synthesizing enzymes, cell debris, and volatile substances (Bhattacacharyya & Ghosh, 1971; Burger et al., 2004; Campagna et al., 2012). Secretion content varies according to sex, age, diet, and sexual period (Sandilands et al., 2004; Serbest, 2004; Zık & Erdost, 2002).
Although the function of the gland is controversial, its secretion has been reported to have effects such as feather care (Chiale et al., 2021; Reynolds, 2013), indirect cell regeneration, cell growth, and differentiation (Liu, Qi, et al., 2022), antibacterial (Chiale et al., 2020; Galván et al., 2008; Shawkey et al., 2003), pheromone-like in sexual behavior (Liu et al., 2022a; Zhang et al., 2010), protection against predators, or parasites (Caspers et al., 2021).
Histologically, the gland is surrounded externally by a dense connective tissue capsule (Carril et al., 2019; Chiale et al., 2016). Gland epithelium consists of different cellular layers in the form of a germinative layer, intermediate layer, secretory layer, and degenerative layer (Carril et al., 2019; Jacob & Zeman, 1972). Some studies have reported that the gland is a compound tubulo-alveolar holocrine, ectodermal invagination (Jacob & Ziswiler, 1982; Sandilands et al., 2004; Wagner & Boord, 1975). In addition, it has been reported that the gland in quail, roosters, Aseel breed roosters and chickens, and ducks has a simple tubular structure with holocrine-type secretion (Çınar & Kelek, 2014; Harem et al., 2005; Suzuki & Kusuhara, 1996; Yılmaz et al., 2018; Zık & Erdost, 2002).
The turkey (Meleagris gallopavo) is a bird in the order Galliformes, family Phasanidae. It has been recognized as a species with several subspecies of its wild ancestor spread over a wide area from southern Canada to southern Mexico. It was reported to have been domesticated before Christopher Columbus discovered the Americas (Crawford, 1992; Thornton, 2016). The structure of the uropygial gland in some bird species in the order Galliformes has been studied macroscopically and microscopically (Çınar & Kelek, 2014; Demırcıoglu et al., 2019; Rajathi, 2019). Apart from routine triple and Hematoxylin & Eosin (H&E) stains, histochemical stains are available (Chiale et al., 2016, 2019; Çınar & Kelek, 2014). In addition, scanning electron microscopy (SEM) studies are quite limited. In the literature, there is a study on macroscopic and some morphometric features of the uropygial gland in turkeys (Rajathi, 2019). However, no macroscopic, morphometric, or microscopic examination of this gland about sex differences was found. This study is the first study to investigate macroscopically, morphometrically, and microscopically the uropygial gland in turkeys depending on sex. Microscopic examination was performed using light and SEM.
Glandulae uropygialis of a total of 13 turkeys (7 females and 6 males) were used as material in the study. The glands were obtained from turkeys that did not show any clinical findings and died from reasons unrelated to the study. The tissues were delivered fresh to the laboratory. The turkeys were brought to the laboratory in autumn and no evidence of a reproductive cycle was observed.
In our study, the presence of the uropygial gland was observed in all turkeys. The gland was topographically located on the last caudal vertebrae. It was covered with a thin skin (Figure 2a). The gland was observed to be embedded in a layer of adipose tissue (Figure 2b). It was determined that the gland consisted of two lobes and a papilla system (Figure 2c,d). The lobes had a symmetrical appearance from an external view. The dorsal and ventral surfaces showed a distinct convexity. It was observed that it resembled the heart in shape and the ends of the lobes merged caudally. There was a short papilla at the caudal end of the gland. Numerous holes (porus glandulae uropygialis) were observed on the papilla, which allowed the secretion to drain out. In addition, many feather structures (Figure 2c,d) were observed in these holes. The gland was divided into two lobes by a septum interlobare (Figure 3). In addition, the presence of ducts named ductus glandulae uropygialis was determined under a stereomicroscope (Figure 3). Inside the lobes, a brown secretion material with a dense consistency attracted attention. The general characterization of glandula uropygialis in turkeys was presented in Table 1.
In male and female turkeys, uropygial gland weights were measured as 2.50 ± 0.16 g and 3.04 ± 0.21 g, respectively. As a result of the statistical evaluation, glandula uropygialis lobe length, papilla width, and papilla height (p < .05) parameters were significantly different between the sexes. It was determined that glandula uropygialis lobe width, papilla width, and papilla height parameters were larger in male turkeys than in females. At the same time, as a result of the index calculations, it was determined that there was no difference between the sexes in terms RGWI, LGUI, and GULI, while PUI value was higher in male turkeys and showed a statistical difference between the sexes (p < .05). Descriptive statistical values and p values of measurement parameters and index values are presented in Tables 2 and 3, respectively.
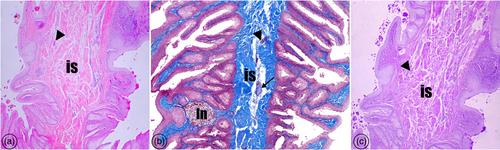
The first histological finding was that the glandula uropygialis was embedded in a dense adipose tissue (Figure 4). In each of the histological staining, both female and male turkeys' uropygial glands histological structures were shown the same structures. The gland was a bi-lobed structure surrounded by a thick fibrous capsule. There were blood and lymph vessels, nerve fibers, adipocytes, and smooth muscle bands in the capsule. The capsule was divided the gland ventrally down between the lobes to form the interlobular septum where blood and lymph vessels, nerve fibers, and adipocytes were located (Figures 5 and 6). The gland structure was determined to be tubular. It was seen to have a simple branched tubular structure in some areas, but mostly a simple tubular gland structure. Each lobe of the organ consisted of several secretory tubules. All tubules were separated from each other by a thin connective tissue. The secretory tubules were divided into central and peripheral tubules. Although the lumens of the central tubules were wide, no distinct lumen structure was observed in the peripheral tubules (Figure 7). Secretory products were observed in some of the central tubule's lumen (Figures 7 and 8). The central and peripheral secretory tubules were composed of four distinct layers differing in shape and cell arrangement basal cell layer, intermediate cell layer, secretory cell layer, and degenerative cell layer (Figure 8). The intermediate cell layer consisted of 2–3 layers of polyhedral cells with eosinophilic cytoplasm and centrally located nuclei. Different-sized, vacuolated, polyhedral cells, and containing secretory granules, were observed to form the secretory cell layer. The degenerated cell layer adjacent to the tubule lumen was composed of cells with pyknotic nuclei and pale cytoplasm (Figure 8). In the parts of the interlobular septum close to the papilla, dense amounts of adipose tissue, nerve fibers, and smooth muscle structures were observed. In our study, lymphoid tissue cell communities were located both between the tubular glands (Figure 7a) and in the connective tissue forming the interlobular septum (Figure 6b). The excrete ducts opened into two big collector ducts (ductus glandulae uropygialis) caudal to the gland (Figure 9a,b). PAS (+) reaction was observed in the basal membrane and secretory material (Figure 8b). The fact that the reaction was not glycogen-induced was determined by the observation of PAS positivity as a result of staining with the PAS-diastase technique (Figure 10a). As a result of Orcein staining, elastic fibers were found in the capsule surrounding the uropygial gland and in the intertubular and interlobular septum (Figure 10b).
SEM revealed the presence of a thick connective tissue capsule outside the uropygial gland (Figure 11b,c). Glandular lobes consisted of lobules variable in size and shape (Figure 11). The lobules consisted of different shaped follicles (sphere-like) protruding from the surface of the gland and marked by a thick band of connective tissue on the outside (Figure 12a,b). It was observed that these glandular follicle structures were gathered together in different shapes or mostly disorganized. It was also observed that these follicle structures were connected to each other by connective tissue threads (interfollicular septa) (Figure 12a). The duct opening of each lobe was separated from the gland surface by a shallow groove containing a mass of lipid secretion radiating from the gland surface towards the ductal wall. This opening was observed to be oval in shape, of varying thickness and with a folded edge (Figure 12c). Multiple holes (porus glandulae uropygialis) were observed caudal to the papilla (Figure 12d).
Although the glandula uropygialis is present in almost all bird species during the embryonic period, it may be vestigialised in adults of some orders, families, genera, and species (Johnston, 1988; Salibian & Montalti, 2009). In our study, uropygial gland was present in all turkeys examined.
It has been reported that the gland is located in the region between the fourth caudal vertebra and the pygostyle in different bird species (Lucas & Stettenheim, 1972; Yılmaz et al., 2018), at the base of the tail, on the pygostyle muscles. (Johnston, 1988; Sawad, 2006) Between the caudal side of the lumbosacral bone and the first coccygeal vertebra (Kozlu et al., 2011) and generally on the free caudal vertebrae (Moreno-Rueda, 2016; Yılmaz & Yılmaz, 2019). Our study determined that the uropygial gland was located on the last caudal vertebra in the literature.
Similar to Rajathi (2019) and Lucas & Stettenheim, 1972 (fowl and quail), it was determined as a two-lobed gland. The uropygial gland in this study had two lobes, right and left, and a common papilla with ducts draining secretion from these glands. The lobes were orientated forwards and outwards. The lobes of the uropygial gland were pear-shaped, as in turkeys and chickens (Farner et al., 1982; Rajathi, 2019).
The common papilla of the gland was thick and elongated as reported in domestic chickens (Lucas & Stettenheim, 1972; Rajathi, 2019). The papillae were surrounded by skin covered with small feathers extending dorsocaudally, as in chickens (Lucas & Stettenheim, 1972).
In domestic chickens, it has been reported that the ducts open outwards with a breast-like papilla located in the midline, caudal, and dorsal (Gezici, 2002). The papilla structure was reported to be conical in Psittaciformes species and cylindrical in Monk parrots (Carril et al., 2019). In this study, the papilla structure of the gland was determined to be single, thick, and long, similar to domestic chickens (Lucas & Stettenheim, 1972) and turkeys (Rajathi, 2019). In our study, it was determined that the papilla had two separate main ducts coming from each lobe. Similar results have been found in poultry and Japanese quail (Farner et al., 1982; Mobini & Ziaii, 2011; Rajathi, 2019). In addition, similar to studies in chickens and roosters (Yılmaz & Yılmaz, 2019), it was found that numerous feather structures surrounded the papilla. Numerous holes (porus glandulae uropygialis) were observed at the caudal end of the papilla structure. These holes allow the secretion of secretion from the two uropygial gland ducts.
Age, season, and sex have been found to have a significant effect on the weight of the uropygial gland (Chen et al., 2015; Elder, 1954). In our study, the weight of uropygial glands in male and female turkeys was found to be 2.50 ± 0.16 g and 3.04 ± 0.21 g, respectively. The same value was reported as 0.95 ± 0.15 and 0.91 ± 0.26 g in Aseel roosters and chicks, respectively (Yılmaz et al., 2018). Demırcıoglu et al. (2019) reported the relative gland weight as 0.11 ± 0.01 g in quails. The average value of RGWI in Aseel breed hens and roosters was found to be 0.0423 (Yılmaz et al., 2018) and 0.094 ± 0.041 in Gallus gallus (Montalti et al., 2000). Rajathi (2019) determined this index value as 0.01 ± 0.01 in turkeys, while 0.03 ± 0.001 was found in our study. There was no statistical difference between male and female animals in terms of RGWI value.
Rajathi (2019) reported right and left lobe length parameters in turkeys as 9.7 ± 0.2 mm and 9.6 ± 0.2 mm, respectively. In a study conducted in quails, right GUL was determined as 10.31 mm in females and 10.39 mm in males (Demırcıoglu et al., 2019). In our study, GUL parameter was found to be higher than the values determined by Rajathi. In addition, while there was a statistical difference between male and female animals, the value was higher in male animals. Similarly, PUH and PUW parameters had a statistically significant difference between male and female animals in our study. PUH value was higher in quail (Demırcıoglu et al., 2019), Aseel breed roosters and hens (Yılmaz et al., 2018) and turkeys (Rajathi, 2019). In our study, it was determined that these parameters were greater in male turkeys than females.
Histologic examination revealed that the glandula uropygialis was surrounded by a capsule consisting of a thick connective tissue layer on the outside, similar to studies in quail, partridge, and chickens. There are two lobes and a papilla system under the capsule (Demırcıoglu et al., 2019; Önal et al., 2013; Yılmaz & Yılmaz, 2020). The capsule structure contained blood and lymph vessels, nerve fibers, adipocytes, and smooth muscle bands. Kozlu et al. (2011) and Chiale et al. (2016) reported that the excretory ducts opening the glands were located in the center of the lobe. In our study, similar to the report of Yılmaz and Yılmaz (2020), ducts opened into two main excretory ducts (ductus glandulae uropygialis) caudal to the gland. It is consistent with the literature that the secretory tubules of both lobes are divided into central and peripheral tubules and located around a central lumen (Balkaya et al., 2016; Kozlu et al., 2011; Önal et al., 2013; Stangier et al., 2023). Contrary to the studies reporting that the gland has a compound tubulo-alveolar holocrine character (Jacob & Ziswiler, 1982; Sandilands et al., 2004; Wagner & Boord, 1975), in our study, it was determined that it has a simple tubular structure with holocrine secretion as reported in many studies (Çınar & Kelek, 2014; Harem et al., 2005; Suzuki & Kusuhara, 1996; Yılmaz et al., 2018; Zık & Erdost, 2002).
Similar to studies in quails and guinea fowl, in our study, central and peripheral secretory tubules were found to consist of four different cell layers differing in shape and cell arrangement from periphery to center: basal cell layer, intermediate cell layer, secretory cell layer, and degenerative cell layer (Ushakumary et al., 2011; Zık & Erdost, 2002). Although the lumens of the central tubules were wide, no distinct lumen structure was observed in the peripheral tubules.
The presence of smooth muscle cells, blood, and lymph vessels in the structure of the gland, especially in the septum interlobare and intertubular septum, has been reported (Yılmaz et al., 2018). Similarly, smooth muscle cells were found especially in trabeculae in magpies (Balkaya et al., 2016). However, some studies have reported that the glandula uropygialis of poultry species such as geese (Hou, 1928), grouse (Sawad, 2006), and starlings (Sadoon, 2011) are completely devoid of smooth muscle cells. In our study, smooth muscle cells were observed in the regions of the interlobular septum close to the papilla. It was determined that the intertubular septum was composed of connective tissue and did not contain smooth muscle. Blood and lymph vessels, nerve fibers, and adipocytes were also observed in the interlobular septum.
In a study in Pekin ducks, it was reported that small basal cells with flat nuclei located on the basement membrane, large cells filled with lipid droplets towards the lumen of the tubules and cells with pyknotic nuclei and large lipid droplets close to the lumen were found in the gland (Carpenter & Goodridge, 1988). In roosters, it was determined as flat nuclei with acidophilic cytoplasm located on the basement membrane (Zık & Erdost, 2002). In our study, the basal cell layer consisted of cells with darkly stained nuclei and basophilic cytoplasm. Yılmaz et al. (2018) reported that intermediate cells have acidophilic cytoplasm and basophilic nuclei, secretory cells have many large and small, white-colored fat vacuoles in the lumen, while degenerative cells have larger fat vacuoles in the cytoplasm. In our study, the intermediate cell layer consisted of 2–3 rows of centrally located polyhedral cells with eosinophilic cytoplasm. The secretory cell layer was composed of polyhedral cells of different sizes with vacuoles and secretory granules. The degenerated cell layer adjacent to the tubule lumen consisted of cells with pyknotic nuclei and pale cytoplasm. Acidophilic cells were also detected in H&E staining. The high number of acidophilic cells indicates that there is a large amount of fat synthesis in the gland (Yılmaz et al., 2018). Herbst corpuscles, which were first identified in wild ducks (Harem et al., 2005), were also determined in laughing pigeons (Madkour et al., 2023). Similar to the study in mallard ducks (Yılmaz & Yılmaz, 2019), Herbst corpuscles structure was not detected in our study.
In a study on the rooster uropygial gland, the apical half of the tubules, which were found to be rich in glycogen by the PAS staining method, was named as glycogen region and it was reported that this region gave a strong PAS (+) reaction (Zık & Erdost, 2002). In the female and male partridge uropygial gland, no PAS (+) reaction was observed in the sebaceous region, while in females, it was determined that neutral mucosubstance reacted with moderate intensity in the peripheral parts of some tubules belonging to the sebaceous region. In addition, a strong PAS reaction was detected in females and a moderate to strong PAS reaction was detected in males in the inter-tubular area of the fat region (Önal et al., 2013). In Chilean Flamingo and Monk parakeets, a positive reaction in the uropygial glands with PAS and Alcian Blue (pH 2.5) revealed the presence of glycoconjugates containing carboxyl groups and sulfated esters. These compounds have been associated with protective functions (protection and maintenance of feathers) in digestive tract organs in fish (Díaz et al., 2008; Yashpal et al., 2014) and uropygial glands in birds (Carril et al., 2019; Chiale et al., 2021). In our study, positive staining with PAS was observed especially in the basement membrane and secretory material. After PAS diastase staining, similar to Chiale et al. (2016), it was determined that the staining was not glycogen-induced. As a result of Orcein staining, elastic fibers were observed in the capsule and intertubular septum, which is consistent with the literature (Carril et al., 2019; Chiale et al., 2016, 2020, 2021).
Studies have reported the presence of lymphoid cell communities between tubular glands (Harem et al., 2005; Sawad, 2006; Shafiian & Mobini, 2014). It has also been reported that lymphoid cell communities are located in the connective tissue and papillae at the junction of the glands and excretory duct (Chiale et al., 2016; Yılmaz & Yılmaz, 2020). In our study, it was determined that lymphoid tissue cell communities were located both between tubular glands and in connective tissue. Also contains abundant dense connective tissue, smooth muscle fibers, and lymphoid tissue.
Madkour et al., 2023 reported that the glandular lobe consisted of elliptical, triangular, round glandular lobules, and glandular follicle structures came together in different shapes as a result of SEM analysis. In our study, glandular lobules were spherical and elliptical, and glandular follicle structures were mostly scattered. While the duct opening was reported to be oval shaped, with curved edges, and symmetrical thickness Madkour et al. (2023), in our study it was found to be oval shaped, of variable thickness, and with curved edges. There was no difference in the histologic and electron microscopic structure of the glandula uropygialis in male and female turkeys used in the study.
Although a study was conducted by Rajathi in 2019 to determine morphometric features in turkeys, this is the first study to perform light and electron microscopic identification of the glandula uropygialis in turkeys. The macroanatomical, morphometric, and microscopic findings of the glandula uropygialis in the turkeys used in the study were found to be very similar to those of most avian species, but different from those of aquatic species. The differences may be due to species and habitat differences.
Fatma İşbilir: Data curation; investigation; validation; formal analysis; supervision; conceptualization; methodology; resources; writing – original draft; software; visualization; project administration; writing – review and editing. Cansel Güzin Özgüden Akkoç: Methodology; formal analysis; supervision; writing – original draft; writing – review and editing; visualization. İhsan İşbilir: Methodology; visualization; writing – original draft; writing – review and editing; formal analysis; supervision. Özgür Özöner: Methodology; supervision; visualization; writing – review and editing; writing – original draft; formal analysis. Barış Can Güzel: Methodology; formal analysis; supervision; writing – original draft; writing – review and editing. İlker ARICAN: Methodology; formal analysis; supervision; writing – original draft; writing – review and editing. Şükrü Hakan Atalgın: Visualization; writing – original draft; writing – review and editing; conceptualization; methodology. Mehmet Can: Conceptualization; methodology; visualization; writing – review and editing; writing – original draft. Soner Bekmezci: Conceptualization; methodology; visualization; writing – review and editing; writing – original draft.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


