Myra F. Laird, Taylor A. Polvadore, Gabrielle A. Hirschkorn, Julie C. McKinney, Callum F. Ross, Andrea B. Taylor, Claire E. Terhune, Jose Iriarte-Diaz
{"title":"Eulemur 和 Varecia 在咬合力和咬合口之间的权衡。","authors":"Myra F. Laird, Taylor A. Polvadore, Gabrielle A. Hirschkorn, Julie C. McKinney, Callum F. Ross, Andrea B. Taylor, Claire E. Terhune, Jose Iriarte-Diaz","doi":"10.1002/jmor.21699","DOIUrl":null,"url":null,"abstract":"<p>In 1974, Sue Herring described the relationship between two important performance variables in the feeding system, bite force and gape. These variables are inversely related, such that, without specific muscular adaptations, most animals cannot produce high bite forces at large gapes for a given sized muscle. Despite the importance of these variables for feeding biomechanics and functional ecology, the paucity of in vivo bite force data in primates has led to bite forces largely being estimated through ex vivo methods. Here, we quantify and compare in vivo bite forces and gapes with output from simulated musculoskeletal models in two craniofacially distinct strepsirrhines: <i>Eulemur</i>, which has a shorter jaw and slower chewing cycle durations relative to jaw length and body mass compared to <i>Varecia</i>. Bite forces were collected across a range of linear gapes from 16 adult lemurs (suborder Strepsirrhini) at the Duke Lemur Center in Durham, North Carolina representing three species: <i>Eulemur flavifrons</i> (<i>n</i> = 6; 3F, 3M), <i>Varecia variegata</i> (<i>n</i> = 5; 3F, 2M), and <i>Varecia rubra</i> (<i>n</i> = 5; 5F). Maximum linear and angular gapes were significantly higher for <i>Varecia</i> compared to <i>Eulemur</i> (<i>p</i> = .01) but there were no significant differences in recorded maximum in vivo bite forces (<i>p</i> = .88). Simulated muscle models using architectural data for these taxa suggest this approach is an accurate method of estimating bite force-gape tradeoffs in addition to variables such as fiber length, fiber operating range, and gapes associated with maximum force. Our in vivo and modeling data suggest <i>Varecia</i> has reduced bite force capacities in favor of absolutely wider gapes compared to <i>Eulemur</i> in relation to their longer jaws. Importantly, our comparisons validate the simulated muscle approach for estimating bite force as a function of gape in extant and fossil primates.</p>","PeriodicalId":16528,"journal":{"name":"Journal of Morphology","volume":"285 5","pages":""},"PeriodicalIF":1.5000,"publicationDate":"2024-05-07","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1002/jmor.21699","citationCount":"0","resultStr":"{\"title\":\"Tradeoffs between bite force and gape in Eulemur and Varecia\",\"authors\":\"Myra F. Laird, Taylor A. Polvadore, Gabrielle A. Hirschkorn, Julie C. McKinney, Callum F. Ross, Andrea B. Taylor, Claire E. Terhune, Jose Iriarte-Diaz\",\"doi\":\"10.1002/jmor.21699\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p>In 1974, Sue Herring described the relationship between two important performance variables in the feeding system, bite force and gape. These variables are inversely related, such that, without specific muscular adaptations, most animals cannot produce high bite forces at large gapes for a given sized muscle. Despite the importance of these variables for feeding biomechanics and functional ecology, the paucity of in vivo bite force data in primates has led to bite forces largely being estimated through ex vivo methods. Here, we quantify and compare in vivo bite forces and gapes with output from simulated musculoskeletal models in two craniofacially distinct strepsirrhines: <i>Eulemur</i>, which has a shorter jaw and slower chewing cycle durations relative to jaw length and body mass compared to <i>Varecia</i>. Bite forces were collected across a range of linear gapes from 16 adult lemurs (suborder Strepsirrhini) at the Duke Lemur Center in Durham, North Carolina representing three species: <i>Eulemur flavifrons</i> (<i>n</i> = 6; 3F, 3M), <i>Varecia variegata</i> (<i>n</i> = 5; 3F, 2M), and <i>Varecia rubra</i> (<i>n</i> = 5; 5F). Maximum linear and angular gapes were significantly higher for <i>Varecia</i> compared to <i>Eulemur</i> (<i>p</i> = .01) but there were no significant differences in recorded maximum in vivo bite forces (<i>p</i> = .88). Simulated muscle models using architectural data for these taxa suggest this approach is an accurate method of estimating bite force-gape tradeoffs in addition to variables such as fiber length, fiber operating range, and gapes associated with maximum force. Our in vivo and modeling data suggest <i>Varecia</i> has reduced bite force capacities in favor of absolutely wider gapes compared to <i>Eulemur</i> in relation to their longer jaws. Importantly, our comparisons validate the simulated muscle approach for estimating bite force as a function of gape in extant and fossil primates.</p>\",\"PeriodicalId\":16528,\"journal\":{\"name\":\"Journal of Morphology\",\"volume\":\"285 5\",\"pages\":\"\"},\"PeriodicalIF\":1.5000,\"publicationDate\":\"2024-05-07\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://onlinelibrary.wiley.com/doi/epdf/10.1002/jmor.21699\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"Journal of Morphology\",\"FirstCategoryId\":\"3\",\"ListUrlMain\":\"https://onlinelibrary.wiley.com/doi/10.1002/jmor.21699\",\"RegionNum\":4,\"RegionCategory\":\"医学\",\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"Q2\",\"JCRName\":\"ANATOMY & MORPHOLOGY\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"Journal of Morphology","FirstCategoryId":"3","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1002/jmor.21699","RegionNum":4,"RegionCategory":"医学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q2","JCRName":"ANATOMY & MORPHOLOGY","Score":null,"Total":0}
Tradeoffs between bite force and gape in Eulemur and Varecia
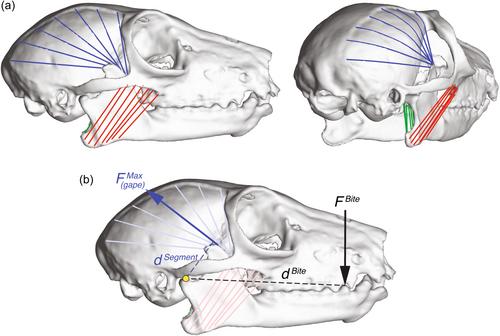
In 1974, Sue Herring described the relationship between two important performance variables in the feeding system, bite force and gape. These variables are inversely related, such that, without specific muscular adaptations, most animals cannot produce high bite forces at large gapes for a given sized muscle. Despite the importance of these variables for feeding biomechanics and functional ecology, the paucity of in vivo bite force data in primates has led to bite forces largely being estimated through ex vivo methods. Here, we quantify and compare in vivo bite forces and gapes with output from simulated musculoskeletal models in two craniofacially distinct strepsirrhines: Eulemur, which has a shorter jaw and slower chewing cycle durations relative to jaw length and body mass compared to Varecia. Bite forces were collected across a range of linear gapes from 16 adult lemurs (suborder Strepsirrhini) at the Duke Lemur Center in Durham, North Carolina representing three species: Eulemur flavifrons (n = 6; 3F, 3M), Varecia variegata (n = 5; 3F, 2M), and Varecia rubra (n = 5; 5F). Maximum linear and angular gapes were significantly higher for Varecia compared to Eulemur (p = .01) but there were no significant differences in recorded maximum in vivo bite forces (p = .88). Simulated muscle models using architectural data for these taxa suggest this approach is an accurate method of estimating bite force-gape tradeoffs in addition to variables such as fiber length, fiber operating range, and gapes associated with maximum force. Our in vivo and modeling data suggest Varecia has reduced bite force capacities in favor of absolutely wider gapes compared to Eulemur in relation to their longer jaws. Importantly, our comparisons validate the simulated muscle approach for estimating bite force as a function of gape in extant and fossil primates.
期刊介绍:
The Journal of Morphology welcomes articles of original research in cytology, protozoology, embryology, and general morphology. Articles generally should not exceed 35 printed pages. Preliminary notices or articles of a purely descriptive morphological or taxonomic nature are not included. No paper which has already been published will be accepted, nor will simultaneous publications elsewhere be allowed.
The Journal of Morphology publishes research in functional, comparative, evolutionary and developmental morphology from vertebrates and invertebrates. Human and veterinary anatomy or paleontology are considered when an explicit connection to neontological animal morphology is presented, and the paper contains relevant information for the community of animal morphologists. Based on our long tradition, we continue to seek publishing the best papers in animal morphology.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


