{"title":"膳食长链脂肪酸代谢可增强抗肿瘤免疫反应。","authors":"Jiaming Wang, Xuetao Cao","doi":"10.1002/cac2.12543","DOIUrl":null,"url":null,"abstract":"<p>Overcoming resistance to immune checkpoint blockade (ICB) therapy will pave the way for effective ICB cancer immunotherapy since a large proportion of cancer patients are not responsive to ICB immunotherapy [<span>1</span>]. The molecular mechanisms of ICB resistance are diverse, including tumor-intrinsic resistant factors (such as genetic and epigenetic disorders), immunosuppressive/disabled factors (such as T cell exhaustion), and environmental restrictive factors (such as neuroendocrine stress and metabolic reprogramming). Deciphering the mechanisms for ICB resistance will provide immense potential for designing new immunotherapeutic strategies in refractory cancers. The tumor microenvironment (TME) includes diverse types of cells, such as immune cells, cancer-associated fibroblasts, and endothelial cells, as well as intercellular cytokines/chemokines, growth factors, and metabolites, which have been recognized as crucial determinants in ICB responsiveness. The recent advent of high-throughput metabolomics and lipidomics analysis reveals that metabolic reprogramming in TME is closely associated with cancer cell invasion, regulated cell death, immune escape, and chemoresistance [<span>2</span>]. On the other hand, immunometabolism also modulates tumor-associated immune cell function and immunotherapy efficacy. For instance, higher expression of major facilitator superfamily domain containing 2A (MFSD2A) in gastric cancer cells inhibits transforming growth factor beta 1 (TGF-β1) production by suppressing cyclooxygenase 2 (COX2)-prostaglandin synthesis, thus promoting antitumor immunity via reprogramming TME [<span>3</span>].</p><p>As a major component of lipids in TME, long-chain fatty acids (LCFAs) are important energy supply and cellular membrane components for cancer cells. Indeed, various types of LCFAs have been found in TME, showing different and sometimes opposite influences on tumor progression and antitumor immunity [<span>4</span>]. For example, palmitic acid promotes metastasis in oral carcinomas and melanoma mouse models through stimulating intratumoral Schwann cells and innervation [<span>5</span>], whereas linoleic acid potentiates CD8<sup>+</sup> T cell antitumor functions via enhancing endoplasmic reticulum-mitochondria contact formation and energetics fitness [<span>6</span>]. Therefore, better understanding of the molecular mechanism for each individual LCFA in tumor immunity is meaningful since it may improve cancer immunotherapy through targeting metabolic reprogramming of TME. In a recent study published in <i>Cell Metabolism</i>, Lai <i>et al.</i> [<span>7</span>] found dietary elaidic acid (EA) supplementation elevates tumoral major histocompatibility complex-1 (MHC-I) expression via acyl-coenzyme A synthetase long chain family member 5 (ACSL5), thus suppressing tumor growth and enhancing anti-programmed cell death protein 1 (anti-PD-1) efficacy (Figure 1).</p><p>Acyl-coenzyme A synthetase long-chain family members (ACSLs) catalyze the conversion of LCFAs into LCFA-coenzyme A (LCFA-CoA) for further lipid catabolism or anabolism. It is reported that interferon gamma (IFNγ) released by cytotoxic T lymphocytes (CTLs) and arachidonic acid in TME coordinately induce tumor cell ferroptosis via acyl-coenzyme A synthetase long chain family member 4 (ACSL4) [<span>8</span>]. Therefore, manipulating ACSLs activity might be an ideal target for metabolic reprogramming in cancer therapy. In this study, the authors found ectopic expression of acyl-coenzyme A synthetase long chain family member 1 (ACSL1), ACSL5, or acyl-coenzyme A synthetase long chain family member (ACSL6) in B16-F10 melanoma cells reduced tumor growth in a subcutaneous mouse model [<span>7</span>]. Given that higher levels of ACSL5 and ACSL6 mRNA were associated with improved overall survival in melanoma patients according to The Cancer Genome Atlas (TCGA) datasets and broad expression of ACSL5 in multiple tissues, the authors focused on ACSL5 for further study. ACSL5 expression also suppressed subcutaneous growth of Lewis lung carcinoma (LLC), Hepa1-6 hepatic tumor, and tumor-model-derived cancer cell line (TDCL) in mouse models. Interestingly, ACSL5-mediated antitumor effect was dependent on acquired immunity since ACSL5 failed to regulate tumor growth in immunodeficient non-obese diabetic-<i>Prkdc</i><sup>scid</sup><i>IL2rg</i><sup>em1</sup>/Smoc (M-NSG) and <i>Rag2</i><sup>−/−</sup> mice or upon anti-CD8 antibody treatment in C57BL/6 mice. Furthermore, ACSL5-deficient TDCL tumors were not responsive to anti-PD-1 therapy in contrast to wide-type tumors, indicating that tumor cell ACSL5 determines the sensitivity to ICB immunotherapy.</p><p>To further delineate ACSL5 antitumor function, the authors examined the number and function of tumor-infiltrating T cells. The number of CD8<sup>+</sup> T cells or CD4 <sup>+</sup> T cells and their cytokines (tumor necrosis factor α [TNF-α] and IFNγ) production were not changed in ACSL5-deficient TDCL tumors. Besides, ACSL5 ectopic B16-F10 tumors had no effects on the number of T cells but slightly increased the TNF-α production by CD8<sup>+</sup> T cells. Although CD8<sup>+</sup> T cell number was increased in ACSL5-expressing LLC tumors, the number of CD4<sup>+</sup> T cells and cytokines production by T cells were not influenced. Therefore, the authors speculated that ACSL5 might regulate tumor sensitivity to T cell-mediated killing. Indeed, they found that ACSL5 deficiency rendered tumor cells more resistant to CD8<sup>+</sup> T cell-mediated cytotoxicity in the co-culture system in vitro. Insufficient tumor immunogenicity is a pivotal cause of ICB resistance since CTLs mediate tumor cell killing through T cell receptor (TCR) recognition of tumor antigen presentation by MHC-I molecules [<span>9</span>]. Accordingly, the authors found downregulation of MHC-I expression in ACSL5-deficient tumor cells. Moreover, knockout of β-2 microglobulin (β2m), a component of MHC-I, promoted tumor growth in control tumors, but not in ACSL5-deficient tumors in vivo, indicating ACSL5 mediates antitumor effect through MHC-I upregulation.</p><p>By transcriptomic analysis of wild-type and ACSL5-deficient cells, the authors noticed that NLR family CARD domain containing 5 (NLRC5), one of the key transcription activators of MHC-I, was reduced upon ACSL5 knockout. Silencing NLRC5 completely abolished the MHC-I upregulation by ACSL5 ectopic expression, suggesting the ACSL5-NLRC5-MHC-I axis in determining tumor cell immunogenicity for induction of antitumor immunity.</p><p>Given that ACSLs take part in LCFAs activation, whether any LCFAs supplementation phenocopies ACSL5's antitumor function? According to various LCFAs screening, the authors found EA, an oleic acid <i>trans</i> isomer and a common component of animal fats and vegetable oils, potently enhanced basal and IFNγ-stimulated MHC-I expression. ACSL5 or NLRC5 deficiency impaired the induction of MHC-I by EA, further validating the EA-ACSL5-NLRC5-MHC-I axis in metabolic regulation of antitumor immunity. Besides, intraperitoneal and intragastrical EA administration both suppressed B16-F10 tumor growth in vivo in an immune-dependent manner. Consistently, EA treatment in combination with anti-PD-1 therapy significantly reduced LLC tumor burden, which was resistant to ICB originally, underscoring the promising therapeutic potential of dietary EA supplementation in promoting responsiveness to ICB. Finally, the authors found that higher plasma EA level was correlated with patients with better response to ICB and improved progression-free survival, implying that plasma EA might be an ideal predictor for ICB effectiveness.</p><p>Although numerous studies suggest the harmful effect of <i>trans</i> fatty acids (TFAs) especially in cardiovascular diseases, this study unveiled an unexpected role of TFAs in boosting tumor immunogenicity and enhancing antitumor immune response. However, there are still many questions that need to be addressed. How the metabolic enzyme ACSL5 regulates nuclear transcription factor NLRC5 expression, and how NLRC5 acts as a fatty acid metabolism alteration sensor are unclear. In contrast to this finding, EA has been reported to promote the growth and metastasis of mouse colorectal cancer cells by activating Wnt and extracellular signal-regulated kinase (ERK) signaling [<span>10</span>]. Whether the opposite functions of EA in different tumor models are due to TME heterogeneity needs further investigation. Although the authors focused on the antitumor effect of EA in tumor cells, whether EA also directly affects CD8<sup>+</sup> T cells function is unclear. It is also interesting to examine the lipid metabolism rewiring, fatty acid-mediated signaling transduction and antitumor function modulation in immune cells during EA intake. The authors demonstrated that ACSL5 converted EA into EA-CoA using isotype labeling, however, whether EA-CoA stimulated MHC-I expression directly through regulating target protein acylation like palmitoylation remains elusive. After conversion to EA-CoA, the activated fatty acids were essential for the downstream lipid metabolic pathways, such as fatty acid β-oxidation (FAO), lipid biosynthesis, and protein acylation (Figure 1). Our unpublished data suggest that ACSL5 can regulate phospholipid synthesis and cellular membrane properties, which might also impact membrane translocation of MHC-1 in tumor cells and other receptors in immune cells. Although the authors excluded the influence of FAO on ACSL5-mediated MHC-I expression in this paper [<span>7</span>], the metabolic perturbation by ACSL5 is still complicated and further exploration of deeper molecular mechanisms underlying EA-ACSL5 axis in tumor immunology is worthwhile.</p><p>Jiaming Wang and Xuetao Cao drafted the manuscript and figures. Xuetao Cao supervised and revised the manuscript.</p><p>The authors declare that they have no competing interests.</p><p>Not applicable.</p>","PeriodicalId":9495,"journal":{"name":"Cancer Communications","volume":"44 5","pages":"580-583"},"PeriodicalIF":20.1000,"publicationDate":"2024-04-12","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1002/cac2.12543","citationCount":"0","resultStr":"{\"title\":\"Dietary long-chain fatty acid metabolism boosts antitumor immune response\",\"authors\":\"Jiaming Wang, Xuetao Cao\",\"doi\":\"10.1002/cac2.12543\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p>Overcoming resistance to immune checkpoint blockade (ICB) therapy will pave the way for effective ICB cancer immunotherapy since a large proportion of cancer patients are not responsive to ICB immunotherapy [<span>1</span>]. The molecular mechanisms of ICB resistance are diverse, including tumor-intrinsic resistant factors (such as genetic and epigenetic disorders), immunosuppressive/disabled factors (such as T cell exhaustion), and environmental restrictive factors (such as neuroendocrine stress and metabolic reprogramming). Deciphering the mechanisms for ICB resistance will provide immense potential for designing new immunotherapeutic strategies in refractory cancers. The tumor microenvironment (TME) includes diverse types of cells, such as immune cells, cancer-associated fibroblasts, and endothelial cells, as well as intercellular cytokines/chemokines, growth factors, and metabolites, which have been recognized as crucial determinants in ICB responsiveness. The recent advent of high-throughput metabolomics and lipidomics analysis reveals that metabolic reprogramming in TME is closely associated with cancer cell invasion, regulated cell death, immune escape, and chemoresistance [<span>2</span>]. On the other hand, immunometabolism also modulates tumor-associated immune cell function and immunotherapy efficacy. For instance, higher expression of major facilitator superfamily domain containing 2A (MFSD2A) in gastric cancer cells inhibits transforming growth factor beta 1 (TGF-β1) production by suppressing cyclooxygenase 2 (COX2)-prostaglandin synthesis, thus promoting antitumor immunity via reprogramming TME [<span>3</span>].</p><p>As a major component of lipids in TME, long-chain fatty acids (LCFAs) are important energy supply and cellular membrane components for cancer cells. Indeed, various types of LCFAs have been found in TME, showing different and sometimes opposite influences on tumor progression and antitumor immunity [<span>4</span>]. For example, palmitic acid promotes metastasis in oral carcinomas and melanoma mouse models through stimulating intratumoral Schwann cells and innervation [<span>5</span>], whereas linoleic acid potentiates CD8<sup>+</sup> T cell antitumor functions via enhancing endoplasmic reticulum-mitochondria contact formation and energetics fitness [<span>6</span>]. Therefore, better understanding of the molecular mechanism for each individual LCFA in tumor immunity is meaningful since it may improve cancer immunotherapy through targeting metabolic reprogramming of TME. In a recent study published in <i>Cell Metabolism</i>, Lai <i>et al.</i> [<span>7</span>] found dietary elaidic acid (EA) supplementation elevates tumoral major histocompatibility complex-1 (MHC-I) expression via acyl-coenzyme A synthetase long chain family member 5 (ACSL5), thus suppressing tumor growth and enhancing anti-programmed cell death protein 1 (anti-PD-1) efficacy (Figure 1).</p><p>Acyl-coenzyme A synthetase long-chain family members (ACSLs) catalyze the conversion of LCFAs into LCFA-coenzyme A (LCFA-CoA) for further lipid catabolism or anabolism. It is reported that interferon gamma (IFNγ) released by cytotoxic T lymphocytes (CTLs) and arachidonic acid in TME coordinately induce tumor cell ferroptosis via acyl-coenzyme A synthetase long chain family member 4 (ACSL4) [<span>8</span>]. Therefore, manipulating ACSLs activity might be an ideal target for metabolic reprogramming in cancer therapy. In this study, the authors found ectopic expression of acyl-coenzyme A synthetase long chain family member 1 (ACSL1), ACSL5, or acyl-coenzyme A synthetase long chain family member (ACSL6) in B16-F10 melanoma cells reduced tumor growth in a subcutaneous mouse model [<span>7</span>]. Given that higher levels of ACSL5 and ACSL6 mRNA were associated with improved overall survival in melanoma patients according to The Cancer Genome Atlas (TCGA) datasets and broad expression of ACSL5 in multiple tissues, the authors focused on ACSL5 for further study. ACSL5 expression also suppressed subcutaneous growth of Lewis lung carcinoma (LLC), Hepa1-6 hepatic tumor, and tumor-model-derived cancer cell line (TDCL) in mouse models. Interestingly, ACSL5-mediated antitumor effect was dependent on acquired immunity since ACSL5 failed to regulate tumor growth in immunodeficient non-obese diabetic-<i>Prkdc</i><sup>scid</sup><i>IL2rg</i><sup>em1</sup>/Smoc (M-NSG) and <i>Rag2</i><sup>−/−</sup> mice or upon anti-CD8 antibody treatment in C57BL/6 mice. Furthermore, ACSL5-deficient TDCL tumors were not responsive to anti-PD-1 therapy in contrast to wide-type tumors, indicating that tumor cell ACSL5 determines the sensitivity to ICB immunotherapy.</p><p>To further delineate ACSL5 antitumor function, the authors examined the number and function of tumor-infiltrating T cells. The number of CD8<sup>+</sup> T cells or CD4 <sup>+</sup> T cells and their cytokines (tumor necrosis factor α [TNF-α] and IFNγ) production were not changed in ACSL5-deficient TDCL tumors. Besides, ACSL5 ectopic B16-F10 tumors had no effects on the number of T cells but slightly increased the TNF-α production by CD8<sup>+</sup> T cells. Although CD8<sup>+</sup> T cell number was increased in ACSL5-expressing LLC tumors, the number of CD4<sup>+</sup> T cells and cytokines production by T cells were not influenced. Therefore, the authors speculated that ACSL5 might regulate tumor sensitivity to T cell-mediated killing. Indeed, they found that ACSL5 deficiency rendered tumor cells more resistant to CD8<sup>+</sup> T cell-mediated cytotoxicity in the co-culture system in vitro. Insufficient tumor immunogenicity is a pivotal cause of ICB resistance since CTLs mediate tumor cell killing through T cell receptor (TCR) recognition of tumor antigen presentation by MHC-I molecules [<span>9</span>]. Accordingly, the authors found downregulation of MHC-I expression in ACSL5-deficient tumor cells. Moreover, knockout of β-2 microglobulin (β2m), a component of MHC-I, promoted tumor growth in control tumors, but not in ACSL5-deficient tumors in vivo, indicating ACSL5 mediates antitumor effect through MHC-I upregulation.</p><p>By transcriptomic analysis of wild-type and ACSL5-deficient cells, the authors noticed that NLR family CARD domain containing 5 (NLRC5), one of the key transcription activators of MHC-I, was reduced upon ACSL5 knockout. Silencing NLRC5 completely abolished the MHC-I upregulation by ACSL5 ectopic expression, suggesting the ACSL5-NLRC5-MHC-I axis in determining tumor cell immunogenicity for induction of antitumor immunity.</p><p>Given that ACSLs take part in LCFAs activation, whether any LCFAs supplementation phenocopies ACSL5's antitumor function? According to various LCFAs screening, the authors found EA, an oleic acid <i>trans</i> isomer and a common component of animal fats and vegetable oils, potently enhanced basal and IFNγ-stimulated MHC-I expression. ACSL5 or NLRC5 deficiency impaired the induction of MHC-I by EA, further validating the EA-ACSL5-NLRC5-MHC-I axis in metabolic regulation of antitumor immunity. Besides, intraperitoneal and intragastrical EA administration both suppressed B16-F10 tumor growth in vivo in an immune-dependent manner. Consistently, EA treatment in combination with anti-PD-1 therapy significantly reduced LLC tumor burden, which was resistant to ICB originally, underscoring the promising therapeutic potential of dietary EA supplementation in promoting responsiveness to ICB. Finally, the authors found that higher plasma EA level was correlated with patients with better response to ICB and improved progression-free survival, implying that plasma EA might be an ideal predictor for ICB effectiveness.</p><p>Although numerous studies suggest the harmful effect of <i>trans</i> fatty acids (TFAs) especially in cardiovascular diseases, this study unveiled an unexpected role of TFAs in boosting tumor immunogenicity and enhancing antitumor immune response. However, there are still many questions that need to be addressed. How the metabolic enzyme ACSL5 regulates nuclear transcription factor NLRC5 expression, and how NLRC5 acts as a fatty acid metabolism alteration sensor are unclear. In contrast to this finding, EA has been reported to promote the growth and metastasis of mouse colorectal cancer cells by activating Wnt and extracellular signal-regulated kinase (ERK) signaling [<span>10</span>]. Whether the opposite functions of EA in different tumor models are due to TME heterogeneity needs further investigation. Although the authors focused on the antitumor effect of EA in tumor cells, whether EA also directly affects CD8<sup>+</sup> T cells function is unclear. It is also interesting to examine the lipid metabolism rewiring, fatty acid-mediated signaling transduction and antitumor function modulation in immune cells during EA intake. The authors demonstrated that ACSL5 converted EA into EA-CoA using isotype labeling, however, whether EA-CoA stimulated MHC-I expression directly through regulating target protein acylation like palmitoylation remains elusive. After conversion to EA-CoA, the activated fatty acids were essential for the downstream lipid metabolic pathways, such as fatty acid β-oxidation (FAO), lipid biosynthesis, and protein acylation (Figure 1). Our unpublished data suggest that ACSL5 can regulate phospholipid synthesis and cellular membrane properties, which might also impact membrane translocation of MHC-1 in tumor cells and other receptors in immune cells. Although the authors excluded the influence of FAO on ACSL5-mediated MHC-I expression in this paper [<span>7</span>], the metabolic perturbation by ACSL5 is still complicated and further exploration of deeper molecular mechanisms underlying EA-ACSL5 axis in tumor immunology is worthwhile.</p><p>Jiaming Wang and Xuetao Cao drafted the manuscript and figures. Xuetao Cao supervised and revised the manuscript.</p><p>The authors declare that they have no competing interests.</p><p>Not applicable.</p>\",\"PeriodicalId\":9495,\"journal\":{\"name\":\"Cancer Communications\",\"volume\":\"44 5\",\"pages\":\"580-583\"},\"PeriodicalIF\":20.1000,\"publicationDate\":\"2024-04-12\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://onlinelibrary.wiley.com/doi/epdf/10.1002/cac2.12543\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"Cancer Communications\",\"FirstCategoryId\":\"3\",\"ListUrlMain\":\"https://onlinelibrary.wiley.com/doi/10.1002/cac2.12543\",\"RegionNum\":1,\"RegionCategory\":\"医学\",\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"Q1\",\"JCRName\":\"ONCOLOGY\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"Cancer Communications","FirstCategoryId":"3","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1002/cac2.12543","RegionNum":1,"RegionCategory":"医学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"ONCOLOGY","Score":null,"Total":0}
引用次数: 0
摘要
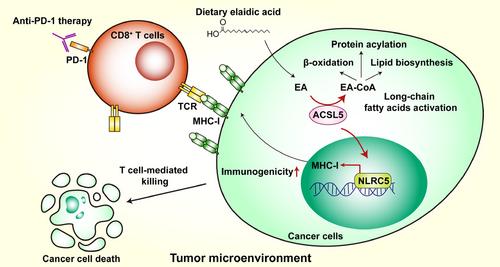
虽然表达ACSL5的LLC肿瘤中CD8+ T细胞数量增加,但CD4+ T细胞数量和T细胞产生的细胞因子不受影响。因此,作者推测 ACSL5 可能会调节肿瘤对 T 细胞介导的杀伤的敏感性。事实上,他们发现,在体外共培养系统中,ACSL5 的缺乏会使肿瘤细胞对 CD8+ T 细胞介导的细胞毒性更具抵抗力。肿瘤免疫原性不足是导致 ICB 抗性的关键原因,因为 CTL 通过 T 细胞受体(TCR)识别 MHC-I 分子呈递的肿瘤抗原来介导肿瘤细胞杀伤 [9]。因此,作者发现 ACSL5 缺失的肿瘤细胞中 MHC-I 表达下调。通过对野生型和 ACSL5 基因缺陷型细胞进行转录组分析,作者注意到,ACSL5 基因敲除后,MHC-I 的关键转录激活因子之一 NLR 家族含 CARD 结构域 5(NLRC5)的表达减少。沉默 NLRC5 可完全消除 ACSL5 异位表达对 MHC-I 的上调,这表明 ACSL5-NLRC5-MHC-I 轴决定了肿瘤细胞的免疫原性,从而诱导抗肿瘤免疫。根据各种 LCFAs 的筛选,作者发现油酸反式异构体 EA(动物脂肪和植物油的常见成分)能有效增强基础和 IFNγ 刺激的 MHC-I 表达。ACSL5或NLRC5缺乏会影响EA对MHC-I的诱导,这进一步验证了EA-ACSL5-NLRC5-MHC-I轴在抗肿瘤免疫代谢调节中的作用。此外,腹腔注射和胃内注射EA都能以免疫依赖的方式抑制体内B16-F10肿瘤的生长。同样,EA与抗PD-1疗法联合使用可显著减少LLC肿瘤负荷,而LLC原本对ICB具有抗药性,这突显了膳食补充EA在促进对ICB的反应性方面具有良好的治疗潜力。最后,作者发现血浆EA水平越高,患者对ICB的反应越好,无进展生存期也越长,这意味着血浆EA可能是ICB疗效的理想预测指标。尽管大量研究表明反式脂肪酸(TFAs)有害,尤其是对心血管疾病有害,但这项研究揭示了反式脂肪酸在增强肿瘤免疫原性和提高抗肿瘤免疫反应方面的意想不到的作用。然而,仍有许多问题需要解决。代谢酶ACSL5如何调控核转录因子NLRC5的表达,以及NLRC5如何作为脂肪酸代谢改变的传感器,这些问题都不清楚。与这一发现相反,有报道称 EA 可通过激活 Wnt 和细胞外信号调节激酶(ERK)信号来促进小鼠结直肠癌细胞的生长和转移[10]。EA在不同肿瘤模型中的相反功能是否是由于TME的异质性造成的,还需要进一步研究。尽管作者关注的是 EA 对肿瘤细胞的抗肿瘤作用,但 EA 是否也直接影响 CD8+ T 细胞的功能尚不清楚。研究摄入 EA 期间免疫细胞的脂质代谢重构、脂肪酸介导的信号转导和抗肿瘤功能调控也很有意义。作者利用同型标记技术证明了 ACSL5 能将 EA 转化为 EA-CoA,然而,EA-CoA 是否直接通过调控靶蛋白酰化(如棕榈酰化)来刺激 MHC-I 的表达仍是未知数。转化为 EA-CoA 后,活化的脂肪酸是下游脂质代谢途径(如脂肪酸 β-氧化(FAO)、脂质生物合成和蛋白质酰化)所必需的(图 1)。我们未发表的数据表明,ACSL5 可调控磷脂合成和细胞膜特性,这也可能影响肿瘤细胞中 MHC-1 和免疫细胞中其他受体的膜转位。虽然作者在本文[7]中排除了FAO对ACSL5介导的MHC-I表达的影响,但ACSL5的代谢扰动仍然复杂,值得进一步探讨EA-ACSL5轴在肿瘤免疫学中的深层分子机制。曹雪涛指导并修改了手稿。作者声明他们没有利益冲突。
Overcoming resistance to immune checkpoint blockade (ICB) therapy will pave the way for effective ICB cancer immunotherapy since a large proportion of cancer patients are not responsive to ICB immunotherapy [1]. The molecular mechanisms of ICB resistance are diverse, including tumor-intrinsic resistant factors (such as genetic and epigenetic disorders), immunosuppressive/disabled factors (such as T cell exhaustion), and environmental restrictive factors (such as neuroendocrine stress and metabolic reprogramming). Deciphering the mechanisms for ICB resistance will provide immense potential for designing new immunotherapeutic strategies in refractory cancers. The tumor microenvironment (TME) includes diverse types of cells, such as immune cells, cancer-associated fibroblasts, and endothelial cells, as well as intercellular cytokines/chemokines, growth factors, and metabolites, which have been recognized as crucial determinants in ICB responsiveness. The recent advent of high-throughput metabolomics and lipidomics analysis reveals that metabolic reprogramming in TME is closely associated with cancer cell invasion, regulated cell death, immune escape, and chemoresistance [2]. On the other hand, immunometabolism also modulates tumor-associated immune cell function and immunotherapy efficacy. For instance, higher expression of major facilitator superfamily domain containing 2A (MFSD2A) in gastric cancer cells inhibits transforming growth factor beta 1 (TGF-β1) production by suppressing cyclooxygenase 2 (COX2)-prostaglandin synthesis, thus promoting antitumor immunity via reprogramming TME [3].
As a major component of lipids in TME, long-chain fatty acids (LCFAs) are important energy supply and cellular membrane components for cancer cells. Indeed, various types of LCFAs have been found in TME, showing different and sometimes opposite influences on tumor progression and antitumor immunity [4]. For example, palmitic acid promotes metastasis in oral carcinomas and melanoma mouse models through stimulating intratumoral Schwann cells and innervation [5], whereas linoleic acid potentiates CD8+ T cell antitumor functions via enhancing endoplasmic reticulum-mitochondria contact formation and energetics fitness [6]. Therefore, better understanding of the molecular mechanism for each individual LCFA in tumor immunity is meaningful since it may improve cancer immunotherapy through targeting metabolic reprogramming of TME. In a recent study published in Cell Metabolism, Lai et al. [7] found dietary elaidic acid (EA) supplementation elevates tumoral major histocompatibility complex-1 (MHC-I) expression via acyl-coenzyme A synthetase long chain family member 5 (ACSL5), thus suppressing tumor growth and enhancing anti-programmed cell death protein 1 (anti-PD-1) efficacy (Figure 1).
Acyl-coenzyme A synthetase long-chain family members (ACSLs) catalyze the conversion of LCFAs into LCFA-coenzyme A (LCFA-CoA) for further lipid catabolism or anabolism. It is reported that interferon gamma (IFNγ) released by cytotoxic T lymphocytes (CTLs) and arachidonic acid in TME coordinately induce tumor cell ferroptosis via acyl-coenzyme A synthetase long chain family member 4 (ACSL4) [8]. Therefore, manipulating ACSLs activity might be an ideal target for metabolic reprogramming in cancer therapy. In this study, the authors found ectopic expression of acyl-coenzyme A synthetase long chain family member 1 (ACSL1), ACSL5, or acyl-coenzyme A synthetase long chain family member (ACSL6) in B16-F10 melanoma cells reduced tumor growth in a subcutaneous mouse model [7]. Given that higher levels of ACSL5 and ACSL6 mRNA were associated with improved overall survival in melanoma patients according to The Cancer Genome Atlas (TCGA) datasets and broad expression of ACSL5 in multiple tissues, the authors focused on ACSL5 for further study. ACSL5 expression also suppressed subcutaneous growth of Lewis lung carcinoma (LLC), Hepa1-6 hepatic tumor, and tumor-model-derived cancer cell line (TDCL) in mouse models. Interestingly, ACSL5-mediated antitumor effect was dependent on acquired immunity since ACSL5 failed to regulate tumor growth in immunodeficient non-obese diabetic-PrkdcscidIL2rgem1/Smoc (M-NSG) and Rag2−/− mice or upon anti-CD8 antibody treatment in C57BL/6 mice. Furthermore, ACSL5-deficient TDCL tumors were not responsive to anti-PD-1 therapy in contrast to wide-type tumors, indicating that tumor cell ACSL5 determines the sensitivity to ICB immunotherapy.
To further delineate ACSL5 antitumor function, the authors examined the number and function of tumor-infiltrating T cells. The number of CD8+ T cells or CD4 + T cells and their cytokines (tumor necrosis factor α [TNF-α] and IFNγ) production were not changed in ACSL5-deficient TDCL tumors. Besides, ACSL5 ectopic B16-F10 tumors had no effects on the number of T cells but slightly increased the TNF-α production by CD8+ T cells. Although CD8+ T cell number was increased in ACSL5-expressing LLC tumors, the number of CD4+ T cells and cytokines production by T cells were not influenced. Therefore, the authors speculated that ACSL5 might regulate tumor sensitivity to T cell-mediated killing. Indeed, they found that ACSL5 deficiency rendered tumor cells more resistant to CD8+ T cell-mediated cytotoxicity in the co-culture system in vitro. Insufficient tumor immunogenicity is a pivotal cause of ICB resistance since CTLs mediate tumor cell killing through T cell receptor (TCR) recognition of tumor antigen presentation by MHC-I molecules [9]. Accordingly, the authors found downregulation of MHC-I expression in ACSL5-deficient tumor cells. Moreover, knockout of β-2 microglobulin (β2m), a component of MHC-I, promoted tumor growth in control tumors, but not in ACSL5-deficient tumors in vivo, indicating ACSL5 mediates antitumor effect through MHC-I upregulation.
By transcriptomic analysis of wild-type and ACSL5-deficient cells, the authors noticed that NLR family CARD domain containing 5 (NLRC5), one of the key transcription activators of MHC-I, was reduced upon ACSL5 knockout. Silencing NLRC5 completely abolished the MHC-I upregulation by ACSL5 ectopic expression, suggesting the ACSL5-NLRC5-MHC-I axis in determining tumor cell immunogenicity for induction of antitumor immunity.
Given that ACSLs take part in LCFAs activation, whether any LCFAs supplementation phenocopies ACSL5's antitumor function? According to various LCFAs screening, the authors found EA, an oleic acid trans isomer and a common component of animal fats and vegetable oils, potently enhanced basal and IFNγ-stimulated MHC-I expression. ACSL5 or NLRC5 deficiency impaired the induction of MHC-I by EA, further validating the EA-ACSL5-NLRC5-MHC-I axis in metabolic regulation of antitumor immunity. Besides, intraperitoneal and intragastrical EA administration both suppressed B16-F10 tumor growth in vivo in an immune-dependent manner. Consistently, EA treatment in combination with anti-PD-1 therapy significantly reduced LLC tumor burden, which was resistant to ICB originally, underscoring the promising therapeutic potential of dietary EA supplementation in promoting responsiveness to ICB. Finally, the authors found that higher plasma EA level was correlated with patients with better response to ICB and improved progression-free survival, implying that plasma EA might be an ideal predictor for ICB effectiveness.
Although numerous studies suggest the harmful effect of trans fatty acids (TFAs) especially in cardiovascular diseases, this study unveiled an unexpected role of TFAs in boosting tumor immunogenicity and enhancing antitumor immune response. However, there are still many questions that need to be addressed. How the metabolic enzyme ACSL5 regulates nuclear transcription factor NLRC5 expression, and how NLRC5 acts as a fatty acid metabolism alteration sensor are unclear. In contrast to this finding, EA has been reported to promote the growth and metastasis of mouse colorectal cancer cells by activating Wnt and extracellular signal-regulated kinase (ERK) signaling [10]. Whether the opposite functions of EA in different tumor models are due to TME heterogeneity needs further investigation. Although the authors focused on the antitumor effect of EA in tumor cells, whether EA also directly affects CD8+ T cells function is unclear. It is also interesting to examine the lipid metabolism rewiring, fatty acid-mediated signaling transduction and antitumor function modulation in immune cells during EA intake. The authors demonstrated that ACSL5 converted EA into EA-CoA using isotype labeling, however, whether EA-CoA stimulated MHC-I expression directly through regulating target protein acylation like palmitoylation remains elusive. After conversion to EA-CoA, the activated fatty acids were essential for the downstream lipid metabolic pathways, such as fatty acid β-oxidation (FAO), lipid biosynthesis, and protein acylation (Figure 1). Our unpublished data suggest that ACSL5 can regulate phospholipid synthesis and cellular membrane properties, which might also impact membrane translocation of MHC-1 in tumor cells and other receptors in immune cells. Although the authors excluded the influence of FAO on ACSL5-mediated MHC-I expression in this paper [7], the metabolic perturbation by ACSL5 is still complicated and further exploration of deeper molecular mechanisms underlying EA-ACSL5 axis in tumor immunology is worthwhile.
Jiaming Wang and Xuetao Cao drafted the manuscript and figures. Xuetao Cao supervised and revised the manuscript.
The authors declare that they have no competing interests.
期刊介绍:
Cancer Communications is an open access, peer-reviewed online journal that encompasses basic, clinical, and translational cancer research. The journal welcomes submissions concerning clinical trials, epidemiology, molecular and cellular biology, and genetics.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


