{"title":"Waking up neural stem cells through inhibition of mitochondrial pyruvate import","authors":"Yajiao Shi, You Wan, Jie Zheng","doi":"10.1002/brx2.13","DOIUrl":null,"url":null,"abstract":"<p>Neurogenesis declines sharply in adulthood, partly because neural stem and progenitor cells (NSPCs) increasingly return to a dormant state as they age. However, innate NSPC pools are preserved in specific brain regions throughout an individual's lifetime. Petrelli et al.<span><sup>1</sup></span> recently reported that inhibiting mitochondrial pyruvate import stimulated NSPCs to transition from a quiescent state to an active state, thereby promoting neurogenesis in both young and middle-aged mice. These findings indicate a novel approach for pro-neurogenic treatments.</p><p>Most neurogenesis in the brain is completed during embryonic development, with only small pools of NSPCs remaining to generate new neurons postnatally. These NSPCs are primarily found in the hippocampal dentate gyrus and the subventricular zone. This biological process, particularly adult hippocampal neurogenesis (AHN), plays a crucial role in specific functions such as pattern separation, learning and memory, and emotional regulation. The pool of NSPCs in the adult brain gradually diminishes but is maintained at a certain level throughout life due to the self-renewal of neural stem cells during symmetric cell division.</p><p>However, the extent of AHN declines much more sharply compared with the age-dependent depletion of the NSC pool. One of the most significant causes for this decline is the increasing rate at which NSCs in the adult brain transition from an active to a dormant state, remaining quiescent and refusing to proliferate to initiate neurogenesis. Thus, activating these dormant quiescent NSCs is pivotal for restoring neurogenesis.</p><p>The mitochondrial pyruvate carrier (MPC) on the inner mitochondrial membrane is responsible for transporting the glycolytic end-product pyruvate from the cytosol into the mitochondria, thereby linking glycolysis to the tricarboxylic acid cycle and oxidative phosphorylation. Given that glycolysis plays a critical role in determining the activity state of NSCs, Petrelli et al.<span><sup>1</sup></span> recently found that MPC expression was highest in quiescent NSPCs, as opposed to those that were active or proliferating.</p><p>Moreover, both pharmacological blockage of MPC in vitro using the specific membrane-penetrating inhibitor UK5099 and selective deletion of the <i>Mpc1</i> gene in NSPCs led to the proliferation of these quiescent NSPCs. Subsequently, the authors aimed to investigate the underlying metabolic mechanism. They ruled out the contribution of lactate elevation resulting from MPC loss-of-function as neither lactate supplementation nor downregulation affected the proliferation of NSPCs.</p><p>Instead, they discovered that the elevated intracellular aspartate, presumed to be due to the upregulation of glutamic-oxaloacetic transaminase activity or enhanced mitochondrial aspartate import, played a significant role in activating quiescent NSPCs. In contrast to quiescent NSPCs, inhibiting MPCs did not affect the proliferation of already active NSPCs or the differentiation of their progeny.</p><p>Lastly, the authors demonstrated that conditional <i>Mpc1</i> deletion in NSPCs increased the number of newly generated neurons without affecting the neurite differentiation in one-month-old mice. Importantly, an increase in the number of newly formed neurons was also observed in middle-aged mice (9–11 months of age) with conditional <i>Mpc1</i> deletion in NSPCs (Figure 1).</p><p>This important study sheds light on the critical role of mitochondrial pyruvate metabolism in regulating the balance between quiescence and activation in NSPCs. It also sparks increased interest in exploring the relationship between metabolism and stem cell biology across various disciplines, including neuroscience, cancer research, and tissue engineering.</p><p>It has been observed that the total pyruvate dehydrogenase complex increases with age in mice brains.<span><sup>2</sup></span> However, since pyruvate-involved oxidative activity and glucose metabolism have different effects on neuronal developmental timing in humans and mice,<span><sup>3</sup></span> it warrants further investigation to determine whether MPC inhibition can also facilitate NSPC activation and neurogenesis in humans. Another question to consider is whether the forced activation of quiescent NSPCs through disrupted pyruvate metabolism might alter the fate of NSPC progeny, shifting them from neurogenesis to astrogliosis, or leading to the accelerated depletion of the adult NSC pool. It has been found that this can result in a temporary increase but ultimately long-term deficits in neurogenesis.<span><sup>4</sup></span>\n </p><p>Although there are numerous challenges, neurogenesis-targeted therapies, either by facilitating innate neurogenesis or engrafting exogenous stem cells, still hold great potential for rejuvenating aging brains and treating neurological disorders characterized by neuron loss or neurodegeneration.<span><sup>5</sup></span> Considering the critical role of pyruvate metabolism in NSPCs for initiating neurogenesis, MPC inhibitors, whether administered alone or in combination with stem cell transplantation, could represent novel therapies for neurological injuries and neurodegenerative diseases.</p><p>\n <b>Yajiao Shi</b> and <b>Jie Zheng</b> conceptualized and wrote the manuscript. <b>You Wan</b> revised the paper.</p><p>The authors declare that they have no conflicts of interest.</p>","PeriodicalId":94303,"journal":{"name":"Brain-X","volume":"1 2","pages":""},"PeriodicalIF":0.0000,"publicationDate":"2023-05-29","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1002/brx2.13","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"Brain-X","FirstCategoryId":"1085","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1002/brx2.13","RegionNum":0,"RegionCategory":null,"ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"","JCRName":"","Score":null,"Total":0}
引用次数: 0
Abstract
Neurogenesis declines sharply in adulthood, partly because neural stem and progenitor cells (NSPCs) increasingly return to a dormant state as they age. However, innate NSPC pools are preserved in specific brain regions throughout an individual's lifetime. Petrelli et al.1 recently reported that inhibiting mitochondrial pyruvate import stimulated NSPCs to transition from a quiescent state to an active state, thereby promoting neurogenesis in both young and middle-aged mice. These findings indicate a novel approach for pro-neurogenic treatments.
Most neurogenesis in the brain is completed during embryonic development, with only small pools of NSPCs remaining to generate new neurons postnatally. These NSPCs are primarily found in the hippocampal dentate gyrus and the subventricular zone. This biological process, particularly adult hippocampal neurogenesis (AHN), plays a crucial role in specific functions such as pattern separation, learning and memory, and emotional regulation. The pool of NSPCs in the adult brain gradually diminishes but is maintained at a certain level throughout life due to the self-renewal of neural stem cells during symmetric cell division.
However, the extent of AHN declines much more sharply compared with the age-dependent depletion of the NSC pool. One of the most significant causes for this decline is the increasing rate at which NSCs in the adult brain transition from an active to a dormant state, remaining quiescent and refusing to proliferate to initiate neurogenesis. Thus, activating these dormant quiescent NSCs is pivotal for restoring neurogenesis.
The mitochondrial pyruvate carrier (MPC) on the inner mitochondrial membrane is responsible for transporting the glycolytic end-product pyruvate from the cytosol into the mitochondria, thereby linking glycolysis to the tricarboxylic acid cycle and oxidative phosphorylation. Given that glycolysis plays a critical role in determining the activity state of NSCs, Petrelli et al.1 recently found that MPC expression was highest in quiescent NSPCs, as opposed to those that were active or proliferating.
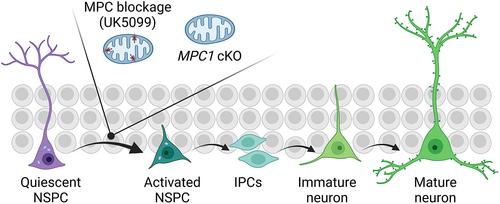
Moreover, both pharmacological blockage of MPC in vitro using the specific membrane-penetrating inhibitor UK5099 and selective deletion of the Mpc1 gene in NSPCs led to the proliferation of these quiescent NSPCs. Subsequently, the authors aimed to investigate the underlying metabolic mechanism. They ruled out the contribution of lactate elevation resulting from MPC loss-of-function as neither lactate supplementation nor downregulation affected the proliferation of NSPCs.
Instead, they discovered that the elevated intracellular aspartate, presumed to be due to the upregulation of glutamic-oxaloacetic transaminase activity or enhanced mitochondrial aspartate import, played a significant role in activating quiescent NSPCs. In contrast to quiescent NSPCs, inhibiting MPCs did not affect the proliferation of already active NSPCs or the differentiation of their progeny.
Lastly, the authors demonstrated that conditional Mpc1 deletion in NSPCs increased the number of newly generated neurons without affecting the neurite differentiation in one-month-old mice. Importantly, an increase in the number of newly formed neurons was also observed in middle-aged mice (9–11 months of age) with conditional Mpc1 deletion in NSPCs (Figure 1).
This important study sheds light on the critical role of mitochondrial pyruvate metabolism in regulating the balance between quiescence and activation in NSPCs. It also sparks increased interest in exploring the relationship between metabolism and stem cell biology across various disciplines, including neuroscience, cancer research, and tissue engineering.
It has been observed that the total pyruvate dehydrogenase complex increases with age in mice brains.2 However, since pyruvate-involved oxidative activity and glucose metabolism have different effects on neuronal developmental timing in humans and mice,3 it warrants further investigation to determine whether MPC inhibition can also facilitate NSPC activation and neurogenesis in humans. Another question to consider is whether the forced activation of quiescent NSPCs through disrupted pyruvate metabolism might alter the fate of NSPC progeny, shifting them from neurogenesis to astrogliosis, or leading to the accelerated depletion of the adult NSC pool. It has been found that this can result in a temporary increase but ultimately long-term deficits in neurogenesis.4
Although there are numerous challenges, neurogenesis-targeted therapies, either by facilitating innate neurogenesis or engrafting exogenous stem cells, still hold great potential for rejuvenating aging brains and treating neurological disorders characterized by neuron loss or neurodegeneration.5 Considering the critical role of pyruvate metabolism in NSPCs for initiating neurogenesis, MPC inhibitors, whether administered alone or in combination with stem cell transplantation, could represent novel therapies for neurological injuries and neurodegenerative diseases.
Yajiao Shi and Jie Zheng conceptualized and wrote the manuscript. You Wan revised the paper.
The authors declare that they have no conflicts of interest.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


