{"title":"Iron-dependent regulation of leaf senescence: a key role for the H2B histone variant HTB4","authors":"Christian Dubos","doi":"10.1111/nph.19199","DOIUrl":null,"url":null,"abstract":"<p>Iron is an essential micronutrient for plant growth and development, as well as for crop productivity and the quality of their derived products (Briat <i>et al</i>., <span>2015</span>). This is because iron is a co-factor for several metalloproteins involved in essential physiological processes such as respiration in mitochondria or photosynthesis in chloroplasts. In most soils, iron is present in the form of insoluble oxides/hydroxides rendering it poorly available to plants. To cope with this poor bioavailability, plants have evolved sophisticated strategies to take up iron from soils (Berger <i>et al</i>., <span>2023</span>; Li <i>et al</i>., <span>2023</span>). Arabidopsis plants preferentially use the reduction-based strategy (the so-called Strategy I), as do most dicots and nongraminous monocots. This strategy relies on the secretion of protons into the rhizosphere by AHA2 (ARABIDOPSIS H<sup>+</sup> ATPASE 2) to decrease the pH of the soil solution and solubilize oxidized iron (Fe<sup>3+</sup>), which is then reduced to Fe<sup>2+</sup> by FRO2 (ferric reduction oxidases 2), and subsequently taken up into the root by IRT1 (iron-regulated transporter 1). This process is tightly regulated since, in excess, iron is also detrimental to the plant because of its capacity to generate reactive oxygen species (ROS) via the Fenton reaction.</p><p>The regulation of Iron homeostasis is well-conserved in plants, and is primarily controlled at the transcriptional level (Berger <i>et al</i>., <span>2023</span>; Li <i>et al</i>., <span>2023</span>). It relies on an intricate regulatory network involving several regulatory proteins, among which the bHLH (basic helix–loop–helix) transcription factors play a preponderant role (Fig. 1). For instance, Arabidopsis has 17 different bHLH proteins (from six bHLH clades) that regulate iron homeostasis. This regulatory network is composed of two modules. The first module relies on FIT/bHLH29 (FER-LIKE IRON DEFICIENCY INDUCED TRANSCRIPTION FACTOR; clade IIIa). FIT is a direct positive regulator of <i>FRO2</i> and <i>IRT1</i> expression (Fig. 1). To achieve its function, FIT interacts with bHLH38, bHLH39, bHLH100, and bHLH101 (clade Ib), forming heterodimers with partially overlapping roles. In the second module, another set of bHLH transcription factors positively regulate the expression of <i>FIT</i> and clade Ib bHLHs (Fig. 1). It involves ILR3/bHLH105 (iaa-leucine resistant 3), IDT1/bHLH34 (iron deficiency tolerant 1), bHLH104, bHLH115 from clade IVc, and URI/bLHL121 (UPSTREAM REGULATOR OF IRT1) from clade IVb (Tissot <i>et al</i>., <span>2019</span>; Gao <i>et al</i>., <span>2020</span>). By contrast, PYE/bHLH47 (POPEYE; clade IVb) is a negative regulator of clade Ib bHLH expression (Pu & Liang, <span>2023</span>).</p><p>In their study, Yang <i>et al</i>. demonstrated that the H2B histone variant HTB4 negatively regulates leaf senescence in an iron-dependent manner. For instance, <i>HTB4</i> expression is induced under iron deficiency and is repressed under iron sufficiency. In addition, <i>htb4</i> loss-of-function mutants display an early leaf senescence phenotype, reduced <i>IRT1</i> and <i>FRO2</i> expression, and decreased iron content. The early leaf senescence phenotype can be reverted by providing extra iron to the <i>htb4</i> mutants, or by overexpressing <i>IRT1</i>. Further investigations provided the molecular mechanism by which HTB4 functions in regulating leaf senescence (Fig. 2). The authors showed that HTB4 promotes the expression of <i>FIT</i> and the four transcription factors of clade Ib bHLH. They further revealed that the binding of HTB4 to the promoter of clade Ib bHLHs correlates with an enrichment of the active mark H3K4me3 (histone H3 lysine 4 tri-methylation) in the vicinity of their transcriptional start sites. As a result, the expression of clade Ib bHLHs is promoted, and in turn, the expression of <i>IRT1</i> and <i>FRO2</i>, fostering root iron uptake and delaying leaf senescence.</p><p>Among the genes whose expression was decreased in the <i>htb4</i> mutant when compared to wild-type plants (transcriptome data analysis), some have been described as playing a key role in regulating iron homeostasis. This is the case for the above-mentioned IDT1 bHLH transcription factor, which is a direct positive regulator of clade Ib bHLHs expression. This is also the case for <i>IMA2</i>/<i>FEP2</i> (<i>IRONMAN 2</i>/<i>FE-UPTAKE-INDUCING PEPTIDE 2</i>) and <i>IMA3</i>/<i>FEP1</i>, which encode peptides that are positive regulators of the iron deficiency response. These peptides inhibit the activity of hemerythrin E3 ubiquitin ligases (i.e. BTS, BRUTUS; BTSL1 and 2, BRUTUS LIKE 1 and 2) that negatively regulate iron homeostasis by promoting the ubiquitin-mediated degradation via the 26S proteasome, of ILR3 and bHLH115 (i.e. BTS), and FIT (i.e. BTSL1 and BTSL2; Selote <i>et al</i>., <span>2015</span>; Rodríguez-Celma <i>et al</i>., <span>2019</span>; Li <i>et al</i>., <span>2021</span>). These observations suggest that HTB4 might also directly regulate the expression of additional genes involved in the transcriptional regulation of iron homeostasis, notably those upstream from FIT and clade Ib bHLHs (Fig. 1). Conversely, it would also be interesting to investigate further if a negative correlation exists between HTB4 and genes that encode proteins that negatively regulate clade Ib expression either directly, such as <i>PYE</i>, or indirectly such as <i>BTS</i>, <i>BTSL1</i>, and <i>BTSL2</i> (Rodríguez-Celma <i>et al</i>., <span>2019</span>; Pu & Liang, <span>2023</span>).</p><p>Yang <i>et al</i>. showed that root iron uptake is impaired in <i>htb4</i> because the expression of <i>IRT1</i> and <i>FRO2</i> was decreased. Whether other mechanisms related to the maintenance of iron homeostasis (e.g. iron translocation, storage, or assimilation) are regulated in an HTB4-dependent manner is also an open question. For instance, the expression of <i>NAS1</i> (<i>nicotianamine synthase 1</i>), <i>NAS4</i> (<i>nicotianamine synthase 4</i>), and <i>OPT3</i> (<i>oligo peptide transporter 3</i>) is reduced in <i>htb4</i> (transcriptome analysis). These three genes participate in the translocation and partitioning of iron between the plant tissues via the phloem, which suggests that these functions are also regulated by HTB4. Moreover, OPT3 is also involved in communicating the shoot iron status to the root to balance the uptake of iron with the plant needs (Mendoza-Cózatl <i>et al</i>., <span>2014</span>).</p><p>At neutral to alkaline pH, iron solubility in soils is low and the activity of ferric reductases, such as FRO2, is strongly impaired (Susin <i>et al</i>., <span>1996</span>). To adapt to such soils, Arabidopsis plants secrete into the rhizosphere iron mobilizing coumarins (i.e. fraxetin; Robe <i>et al</i>., <span>2021a</span>,<span>b</span>). It was recently proposed that once secreted, fraxetin would mostly form Fe<sup>3+</sup>-fraxetin complexes rather than reducing Fe<sup>3+</sup> into Fe<sup>2+</sup>, indicating that the IRT1/FRO2 system does not play a major role in the uptake of iron at alkaline pH. Instead, it is suggested that Fe<sup>3+</sup>-fraxetin complexes are directly taken up into the plant root via a yet unknown mechanism (Robe <i>et al</i>., <span>2021c</span>). Whether or not in such soil environments HTB4 plays a role in modulating iron uptake/homeostasis, and thus leaf senescence, deserves to be investigated.</p><p>Last but not least, it would be of interest to determine if the role of HTB4 in delaying leaf senescence is conserved in grass species such as rice (<i>Oriza sativa</i>) since the uptake of iron mostly relies on an IRT1/FRO2-independent mechanism (Li <i>et al</i>., <span>2021</span>).</p><p>In conclusion, the study of Yang <i>et al</i>. provides a molecular framework by which mineral nutrients such as iron can interfere with leaf senescence. It also opens the road for new investigations to determine the extent of this process either with iron or with other mineral nutrients.</p>","PeriodicalId":48887,"journal":{"name":"New Phytologist","volume":"240 2","pages":"461-463"},"PeriodicalIF":8.1000,"publicationDate":"2023-08-15","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1111/nph.19199","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"New Phytologist","FirstCategoryId":"99","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1111/nph.19199","RegionNum":1,"RegionCategory":"生物学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"Agricultural and Biological Sciences","Score":null,"Total":0}
引用次数: 0
Abstract
Iron is an essential micronutrient for plant growth and development, as well as for crop productivity and the quality of their derived products (Briat et al., 2015). This is because iron is a co-factor for several metalloproteins involved in essential physiological processes such as respiration in mitochondria or photosynthesis in chloroplasts. In most soils, iron is present in the form of insoluble oxides/hydroxides rendering it poorly available to plants. To cope with this poor bioavailability, plants have evolved sophisticated strategies to take up iron from soils (Berger et al., 2023; Li et al., 2023). Arabidopsis plants preferentially use the reduction-based strategy (the so-called Strategy I), as do most dicots and nongraminous monocots. This strategy relies on the secretion of protons into the rhizosphere by AHA2 (ARABIDOPSIS H+ ATPASE 2) to decrease the pH of the soil solution and solubilize oxidized iron (Fe3+), which is then reduced to Fe2+ by FRO2 (ferric reduction oxidases 2), and subsequently taken up into the root by IRT1 (iron-regulated transporter 1). This process is tightly regulated since, in excess, iron is also detrimental to the plant because of its capacity to generate reactive oxygen species (ROS) via the Fenton reaction.
The regulation of Iron homeostasis is well-conserved in plants, and is primarily controlled at the transcriptional level (Berger et al., 2023; Li et al., 2023). It relies on an intricate regulatory network involving several regulatory proteins, among which the bHLH (basic helix–loop–helix) transcription factors play a preponderant role (Fig. 1). For instance, Arabidopsis has 17 different bHLH proteins (from six bHLH clades) that regulate iron homeostasis. This regulatory network is composed of two modules. The first module relies on FIT/bHLH29 (FER-LIKE IRON DEFICIENCY INDUCED TRANSCRIPTION FACTOR; clade IIIa). FIT is a direct positive regulator of FRO2 and IRT1 expression (Fig. 1). To achieve its function, FIT interacts with bHLH38, bHLH39, bHLH100, and bHLH101 (clade Ib), forming heterodimers with partially overlapping roles. In the second module, another set of bHLH transcription factors positively regulate the expression of FIT and clade Ib bHLHs (Fig. 1). It involves ILR3/bHLH105 (iaa-leucine resistant 3), IDT1/bHLH34 (iron deficiency tolerant 1), bHLH104, bHLH115 from clade IVc, and URI/bLHL121 (UPSTREAM REGULATOR OF IRT1) from clade IVb (Tissot et al., 2019; Gao et al., 2020). By contrast, PYE/bHLH47 (POPEYE; clade IVb) is a negative regulator of clade Ib bHLH expression (Pu & Liang, 2023).
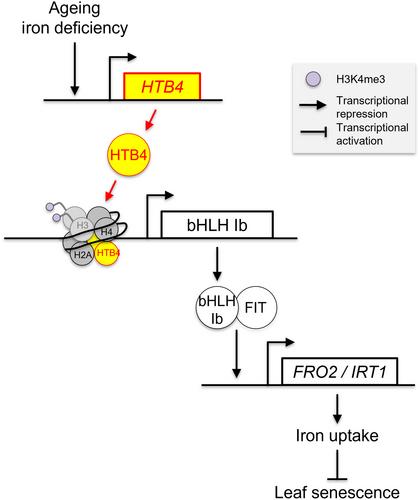
In their study, Yang et al. demonstrated that the H2B histone variant HTB4 negatively regulates leaf senescence in an iron-dependent manner. For instance, HTB4 expression is induced under iron deficiency and is repressed under iron sufficiency. In addition, htb4 loss-of-function mutants display an early leaf senescence phenotype, reduced IRT1 and FRO2 expression, and decreased iron content. The early leaf senescence phenotype can be reverted by providing extra iron to the htb4 mutants, or by overexpressing IRT1. Further investigations provided the molecular mechanism by which HTB4 functions in regulating leaf senescence (Fig. 2). The authors showed that HTB4 promotes the expression of FIT and the four transcription factors of clade Ib bHLH. They further revealed that the binding of HTB4 to the promoter of clade Ib bHLHs correlates with an enrichment of the active mark H3K4me3 (histone H3 lysine 4 tri-methylation) in the vicinity of their transcriptional start sites. As a result, the expression of clade Ib bHLHs is promoted, and in turn, the expression of IRT1 and FRO2, fostering root iron uptake and delaying leaf senescence.
Among the genes whose expression was decreased in the htb4 mutant when compared to wild-type plants (transcriptome data analysis), some have been described as playing a key role in regulating iron homeostasis. This is the case for the above-mentioned IDT1 bHLH transcription factor, which is a direct positive regulator of clade Ib bHLHs expression. This is also the case for IMA2/FEP2 (IRONMAN 2/FE-UPTAKE-INDUCING PEPTIDE 2) and IMA3/FEP1, which encode peptides that are positive regulators of the iron deficiency response. These peptides inhibit the activity of hemerythrin E3 ubiquitin ligases (i.e. BTS, BRUTUS; BTSL1 and 2, BRUTUS LIKE 1 and 2) that negatively regulate iron homeostasis by promoting the ubiquitin-mediated degradation via the 26S proteasome, of ILR3 and bHLH115 (i.e. BTS), and FIT (i.e. BTSL1 and BTSL2; Selote et al., 2015; Rodríguez-Celma et al., 2019; Li et al., 2021). These observations suggest that HTB4 might also directly regulate the expression of additional genes involved in the transcriptional regulation of iron homeostasis, notably those upstream from FIT and clade Ib bHLHs (Fig. 1). Conversely, it would also be interesting to investigate further if a negative correlation exists between HTB4 and genes that encode proteins that negatively regulate clade Ib expression either directly, such as PYE, or indirectly such as BTS, BTSL1, and BTSL2 (Rodríguez-Celma et al., 2019; Pu & Liang, 2023).
Yang et al. showed that root iron uptake is impaired in htb4 because the expression of IRT1 and FRO2 was decreased. Whether other mechanisms related to the maintenance of iron homeostasis (e.g. iron translocation, storage, or assimilation) are regulated in an HTB4-dependent manner is also an open question. For instance, the expression of NAS1 (nicotianamine synthase 1), NAS4 (nicotianamine synthase 4), and OPT3 (oligo peptide transporter 3) is reduced in htb4 (transcriptome analysis). These three genes participate in the translocation and partitioning of iron between the plant tissues via the phloem, which suggests that these functions are also regulated by HTB4. Moreover, OPT3 is also involved in communicating the shoot iron status to the root to balance the uptake of iron with the plant needs (Mendoza-Cózatl et al., 2014).
At neutral to alkaline pH, iron solubility in soils is low and the activity of ferric reductases, such as FRO2, is strongly impaired (Susin et al., 1996). To adapt to such soils, Arabidopsis plants secrete into the rhizosphere iron mobilizing coumarins (i.e. fraxetin; Robe et al., 2021a,b). It was recently proposed that once secreted, fraxetin would mostly form Fe3+-fraxetin complexes rather than reducing Fe3+ into Fe2+, indicating that the IRT1/FRO2 system does not play a major role in the uptake of iron at alkaline pH. Instead, it is suggested that Fe3+-fraxetin complexes are directly taken up into the plant root via a yet unknown mechanism (Robe et al., 2021c). Whether or not in such soil environments HTB4 plays a role in modulating iron uptake/homeostasis, and thus leaf senescence, deserves to be investigated.
Last but not least, it would be of interest to determine if the role of HTB4 in delaying leaf senescence is conserved in grass species such as rice (Oriza sativa) since the uptake of iron mostly relies on an IRT1/FRO2-independent mechanism (Li et al., 2021).
In conclusion, the study of Yang et al. provides a molecular framework by which mineral nutrients such as iron can interfere with leaf senescence. It also opens the road for new investigations to determine the extent of this process either with iron or with other mineral nutrients.
相反,进一步研究HTB4与编码直接(如PYE)或间接(如BTS、BTSL1和BTSL2)负调控分支Ib表达的蛋白质的基因之间是否存在负相关也将是有趣的(Rodríguez-Celma等人,2019;Pu&;Liang,2023)。表明在htb4中根铁吸收受损,因为IRT1和FRO2的表达降低。与维持铁稳态相关的其他机制(如铁转运、储存或同化)是否以HTB4依赖的方式调节也是一个悬而未决的问题。例如,NAS1(烟酰胺合成酶1)、NAS4(烟酰胺合酶4)和OPT3(寡肽转运蛋白3)的表达在htb4中减少(转录组分析)。这三个基因通过韧皮部参与铁在植物组织之间的易位和分配,这表明这些功能也受到HTB4的调节。此外,OPT3还参与将地上部铁的状态传达给根部,以平衡铁的吸收和植物的需求(Mendoza-Cózatl等人,2014)。在中性至碱性pH下,铁在土壤中的溶解度较低,铁还原酶(如FRO2)的活性受到严重损害(Susin等人,1996)。为了适应这种土壤,拟南芥植物分泌到根际调动铁的香豆素(即fraxetin;Robe等人,2021a,b)中。最近有人提出,一旦分泌,fraxetin将主要形成Fe3+-fraxetin复合物,而不是将Fe3+还原为Fe2+,这表明IRT1/FRO2系统在碱性pH下对铁的吸收并不起主要作用。相反,有人认为,Fe3+-fraxetin复合物通过一种未知的机制直接被植物根系吸收(Robe et al.,2021c)。在这种土壤环境中,HTB4是否在调节铁吸收/稳态以及叶片衰老中发挥作用,值得研究。最后但并非最不重要的是,确定HTB4在延缓叶片衰老中的作用是否在水稻(Oriza sativa)等草种中是保守的是有意义的,因为铁的吸收主要依赖于IRT1/FRO2独立机制(Li et al.,2021)。提供了一种分子框架,铁等矿物质营养物质可以通过该分子框架干扰叶片衰老。这也为新的研究开辟了道路,以确定铁或其他矿物营养物质对这一过程的影响程度。
期刊介绍:
New Phytologist is a leading publication that showcases exceptional and groundbreaking research in plant science and its practical applications. With a focus on five distinct sections - Physiology & Development, Environment, Interaction, Evolution, and Transformative Plant Biotechnology - the journal covers a wide array of topics ranging from cellular processes to the impact of global environmental changes. We encourage the use of interdisciplinary approaches, and our content is structured to reflect this. Our journal acknowledges the diverse techniques employed in plant science, including molecular and cell biology, functional genomics, modeling, and system-based approaches, across various subfields.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


