Sharanya Chatterjee, Sujinna Lekmeechai, Nicolas Constantinou, Ewa A. Grzybowska, Zuzanna Kozik, Jyoti S. Choudhary, Cedric N. Berger, Gad Frankel, Abigail Clements
{"title":"The type III secretion system effector EspO of enterohaemorrhagic Escherichia coli inhibits apoptosis through an interaction with HAX-1","authors":"Sharanya Chatterjee, Sujinna Lekmeechai, Nicolas Constantinou, Ewa A. Grzybowska, Zuzanna Kozik, Jyoti S. Choudhary, Cedric N. Berger, Gad Frankel, Abigail Clements","doi":"10.1111/cmi.13366","DOIUrl":null,"url":null,"abstract":"<div>\n \n \n <section>\n \n <p>Many enteric pathogens employ a type III secretion system (T3SS) to translocate effector proteins directly into the host cell cytoplasm, where they subvert signalling pathways of the intestinal epithelium. Here, we report that the anti-apoptotic regulator HS1-associated protein X1 (HAX-1) is an interaction partner of the T3SS effectors EspO of enterohaemorrhagic <i>Escherichia coli</i> (EHEC) and <i>Citrobacter rodentium</i>, OspE of <i>Shigella flexneri</i> and Osp1<sub>STYM</sub> of <i>Salmonella enterica</i> serovar Typhimurium. EspO, OspE and Osp1<sub>STYM</sub> have previously been reported to interact with the focal adhesions protein integrin linked kinase (ILK). We found that EspO localizes both to the focal adhesions (ILK localisation) and mitochondria (HAX-1 localisation), and that increased expression of HAX-1 leads to enhanced mitochondrial localisation of EspO. Ectopic expression of EspO, OspE and Osp1<sub>STYM</sub> protects cells from apoptosis induced by staurosporine and tunicamycin. Depleting cells of HAX-1 indicates that the anti-apoptotic activity of EspO is HAX-1 dependent. Both HAX-1 and ILK were further confirmed as EspO1-interacting proteins during infection using T3SS-delivered EspO1. Using cell detachment as a proxy for cell death we confirmed that T3SS-delivered EspO1 could inhibit cell death induced during EPEC infection, to a similar extent as the anti-apoptotic effector NleH, or treatment with the pan caspase inhibitor z-VAD. In contrast, in cells lacking HAX-1, EspO1 was no longer able to protect against cell detachment, while NleH1 and z-VAD maintained their protective activity. Therefore, during both infection and ectopic expression EspO protects cells from cell death by interacting with HAX-1. These results suggest that despite the differences between EHEC, <i>C</i>. <i>rodentium</i>, <i>Shigella</i> and <i>S</i>. <i>typhimurium</i> infections, hijacking HAX-1 anti-apoptotic signalling is a common strategy to maintain the viability of infected cells.</p>\n </section>\n \n <section>\n \n <h3> Take Away</h3>\n \n <div>\n <ul>\n \n <li>EspO homologues are found in EHEC, <i>Shigella</i>, <i>S</i>. <i>typhimurium</i> and some EPEC.</li>\n \n <li>EspO homologues interact with HAX-1.</li>\n \n <li>EspO protects infected cells from apoptosis.</li>\n \n <li>EspO joins a growing list of T3SS effectors that manipulate cell death pathways.</li>\n </ul>\n </div>\n </section>\n </div>","PeriodicalId":9844,"journal":{"name":"Cellular Microbiology","volume":"23 9","pages":""},"PeriodicalIF":2.6000,"publicationDate":"2021-05-22","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://sci-hub-pdf.com/10.1111/cmi.13366","citationCount":"1","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"Cellular Microbiology","FirstCategoryId":"99","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1111/cmi.13366","RegionNum":2,"RegionCategory":"生物学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q3","JCRName":"CELL BIOLOGY","Score":null,"Total":0}
引用次数: 1
Abstract
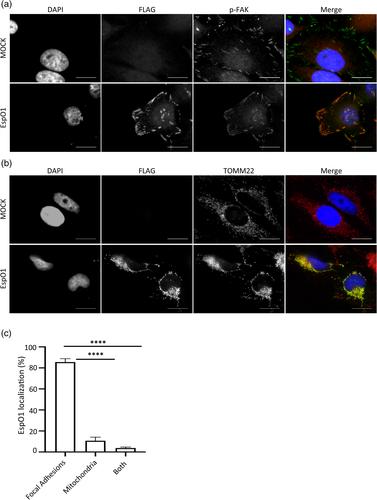
Many enteric pathogens employ a type III secretion system (T3SS) to translocate effector proteins directly into the host cell cytoplasm, where they subvert signalling pathways of the intestinal epithelium. Here, we report that the anti-apoptotic regulator HS1-associated protein X1 (HAX-1) is an interaction partner of the T3SS effectors EspO of enterohaemorrhagic Escherichia coli (EHEC) and Citrobacter rodentium, OspE of Shigella flexneri and Osp1STYM of Salmonella enterica serovar Typhimurium. EspO, OspE and Osp1STYM have previously been reported to interact with the focal adhesions protein integrin linked kinase (ILK). We found that EspO localizes both to the focal adhesions (ILK localisation) and mitochondria (HAX-1 localisation), and that increased expression of HAX-1 leads to enhanced mitochondrial localisation of EspO. Ectopic expression of EspO, OspE and Osp1STYM protects cells from apoptosis induced by staurosporine and tunicamycin. Depleting cells of HAX-1 indicates that the anti-apoptotic activity of EspO is HAX-1 dependent. Both HAX-1 and ILK were further confirmed as EspO1-interacting proteins during infection using T3SS-delivered EspO1. Using cell detachment as a proxy for cell death we confirmed that T3SS-delivered EspO1 could inhibit cell death induced during EPEC infection, to a similar extent as the anti-apoptotic effector NleH, or treatment with the pan caspase inhibitor z-VAD. In contrast, in cells lacking HAX-1, EspO1 was no longer able to protect against cell detachment, while NleH1 and z-VAD maintained their protective activity. Therefore, during both infection and ectopic expression EspO protects cells from cell death by interacting with HAX-1. These results suggest that despite the differences between EHEC, C. rodentium, Shigella and S. typhimurium infections, hijacking HAX-1 anti-apoptotic signalling is a common strategy to maintain the viability of infected cells.
Take Away
EspO homologues are found in EHEC, Shigella, S. typhimurium and some EPEC.
EspO homologues interact with HAX-1.
EspO protects infected cells from apoptosis.
EspO joins a growing list of T3SS effectors that manipulate cell death pathways.
期刊介绍:
Cellular Microbiology aims to publish outstanding contributions to the understanding of interactions between microbes, prokaryotes and eukaryotes, and their host in the context of pathogenic or mutualistic relationships, including co-infections and microbiota. We welcome studies on single cells, animals and plants, and encourage the use of model hosts and organoid cultures. Submission on cell and molecular biological aspects of microbes, such as their intracellular organization or the establishment and maintenance of their architecture in relation to virulence and pathogenicity are also encouraged. Contributions must provide mechanistic insights supported by quantitative data obtained through imaging, cellular, biochemical, structural or genetic approaches.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


