Yuri A Nedialkov, Kristopher Opron, Hailey L Caudill, Fadi Assaf, Amanda J Anderson, Robert I Cukier, Guowei Wei, Zachary F Burton
{"title":"Hinge action versus grip in translocation by RNA polymerase.","authors":"Yuri A Nedialkov, Kristopher Opron, Hailey L Caudill, Fadi Assaf, Amanda J Anderson, Robert I Cukier, Guowei Wei, Zachary F Burton","doi":"10.1080/21541264.2017.1330179","DOIUrl":null,"url":null,"abstract":"<p><p>Based on molecular dynamics simulations and functional studies, a conformational mechanism is posited for forward translocation by RNA polymerase (RNAP). In a simulation of a ternary elongation complex, the clamp and downstream cleft were observed to close. Hinges within the bridge helix and trigger loop supported generation of translocation force against the RNA-DNA hybrid resulting in opening of the furthest upstream i-8 RNA-DNA bp, establishing conditions for RNAP sliding. The β flap tip helix and the most N-terminal β' Zn finger engage the RNA, indicating a path of RNA threading out of the exit channel. Because the β flap tip connects to the RNAP active site through the β subunit double-Ψ-β-barrel and the associated sandwich barrel hybrid motif (also called the flap domain), the RNAP active site is coupled to the RNA exit channel and to the translocation of RNA-DNA. Using an exonuclease III assay to monitor translocation of RNAP elongation complexes, we show that K<sup>+</sup> and Mg<sup>2+</sup> and also an RNA 3'-OH or a 3'-H<sub>2</sub> affect RNAP sliding. Because RNAP grip to template suggests a sticky translocation mechanism, and because grip is enhanced by increasing K<sup>+</sup> and Mg<sup>2+</sup>concentration, biochemical assays are consistent with a conformational change that drives forward translocation as observed in simulations. Mutational analysis of the bridge helix indicates that 778-GARKGL-783 (Escherichia coli numbering) is a homeostatic hinge that undergoes multiple bends to compensate for complex conformational dynamics during phosphodiester bond formation and translocation.</p>","PeriodicalId":47009,"journal":{"name":"Transcription-Austin","volume":"9 1","pages":"1-16"},"PeriodicalIF":4.4000,"publicationDate":"2018-01-01","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://sci-hub-pdf.com/10.1080/21541264.2017.1330179","citationCount":"8","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"Transcription-Austin","FirstCategoryId":"1085","ListUrlMain":"https://doi.org/10.1080/21541264.2017.1330179","RegionNum":0,"RegionCategory":null,"ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"2017/8/30 0:00:00","PubModel":"Epub","JCR":"Q2","JCRName":"BIOCHEMISTRY & MOLECULAR BIOLOGY","Score":null,"Total":0}
引用次数: 8
Abstract
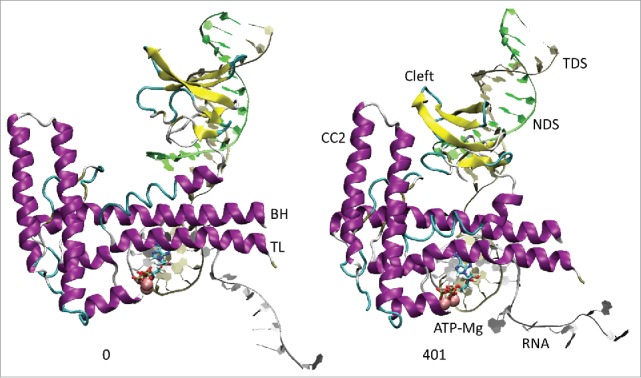
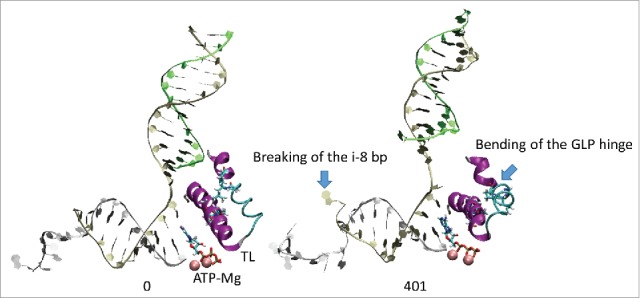
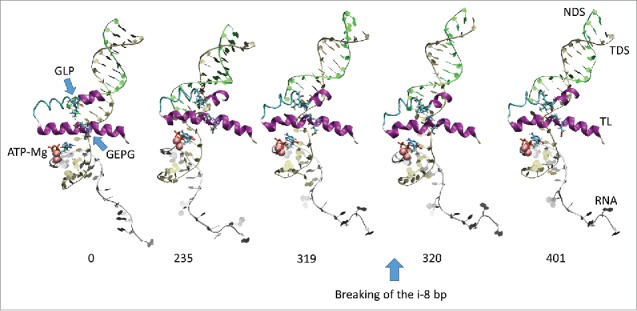
Based on molecular dynamics simulations and functional studies, a conformational mechanism is posited for forward translocation by RNA polymerase (RNAP). In a simulation of a ternary elongation complex, the clamp and downstream cleft were observed to close. Hinges within the bridge helix and trigger loop supported generation of translocation force against the RNA-DNA hybrid resulting in opening of the furthest upstream i-8 RNA-DNA bp, establishing conditions for RNAP sliding. The β flap tip helix and the most N-terminal β' Zn finger engage the RNA, indicating a path of RNA threading out of the exit channel. Because the β flap tip connects to the RNAP active site through the β subunit double-Ψ-β-barrel and the associated sandwich barrel hybrid motif (also called the flap domain), the RNAP active site is coupled to the RNA exit channel and to the translocation of RNA-DNA. Using an exonuclease III assay to monitor translocation of RNAP elongation complexes, we show that K+ and Mg2+ and also an RNA 3'-OH or a 3'-H2 affect RNAP sliding. Because RNAP grip to template suggests a sticky translocation mechanism, and because grip is enhanced by increasing K+ and Mg2+concentration, biochemical assays are consistent with a conformational change that drives forward translocation as observed in simulations. Mutational analysis of the bridge helix indicates that 778-GARKGL-783 (Escherichia coli numbering) is a homeostatic hinge that undergoes multiple bends to compensate for complex conformational dynamics during phosphodiester bond formation and translocation.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


