Chromosome loops arising from intrachromosomal tethering of telomeres occur at high frequency in G1 (non-cycling) mitotic cells: Implications for telomere capture.
{"title":"Chromosome loops arising from intrachromosomal tethering of telomeres occur at high frequency in G1 (non-cycling) mitotic cells: Implications for telomere capture.","authors":"Art Daniel, Luke St Heaps","doi":"10.1186/1475-9268-3-3","DOIUrl":null,"url":null,"abstract":"<p><p>BACKGROUND: To investigate potential mechanisms for telomere capture the spatial arrangement of telomeres and chromosomes was examined in G1 (non-cycling) mitotic cells with diploid or triploid genomes. This was examined firstly by directly labelling the respective short arm (p) and long arm subtelomeres (q) with different fluorophores and probing cell preparations using a number of subtelomere probe pairs, those for chromosomes 1, 3, 4, 5, 6, 7, 9, 10, 12, 17, 18, and 20. In addition some interstitial probes (CEN15, PML and SNRPN on chromosome 15) and whole chromosome paint probes (e.g. WCP12) were jointly hybridised to investigate the co-localization of interphase chromosome domains and tethered subtelomeres. Cells were prepared by omitting exposure to colcemid and hypotonic treatments. RESULTS: In these cells a specific interphase chromosome topology was detected. It was shown that the p and q telomeres of the each chromosome associate frequently (80% pairing) in an intrachromosomal manner, i.e. looped chromosomes with homologues usually widely spaced within the nucleus. This p-q tethering of the telomeres from the one chromosome was observed with large (chromosomes 3, 4, 5), medium sized (6, 7, 9, 10, 12), or small chromosomes (17, 18, 20). When triploid nuclei were probed there were three tetherings of p-q subtelomere signals representing the three widely separated looped chromosome homologues. The separate subtelomere pairings were shown to coincide with separate chromosome domains as defined by the WCP and interstitial probes. The 20% of apparently unpaired subtelomeric signals in diploid nuclei were partially documented to be pairings with the telomeres of other chromosomes. CONCLUSIONS: A topology for telomeres was detected where looped chromosome homologues were present at G1 interphase. These homologues were spatially arranged with respect to one-another independently of other chromosomes, i.e. there was no chromosome order on different sides of the cell nuclei and no segregation into haploid sets was detected. The normal function of this high frequency of intrachromosomal loops is unknown but a potential role is likely in the genesis of telomere captures whether of the intrachromosomal type or between non-homologues. This intrachromosomal tethering of telomeres cannot be related to telomeric or subtelomeric sequences since these are shared in varying degree with other chromosomes. In our view, these intrachromosomal telomeric tetherings with the resulting looped chromosomes arranged in a regular topology must be important to normal cell function since non-cycling cells in G1 are far from quiescent, are in fact metabolically active, and these cells represent the majority status since only a small proportion of cells are normally dividing.</p>","PeriodicalId":84415,"journal":{"name":"Cell & chromosome","volume":"3 1","pages":"3"},"PeriodicalIF":0.0000,"publicationDate":"2004-09-29","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://sci-hub-pdf.com/10.1186/1475-9268-3-3","citationCount":"11","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"Cell & chromosome","FirstCategoryId":"1085","ListUrlMain":"https://doi.org/10.1186/1475-9268-3-3","RegionNum":0,"RegionCategory":null,"ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"","JCRName":"","Score":null,"Total":0}
引用次数: 11
Abstract
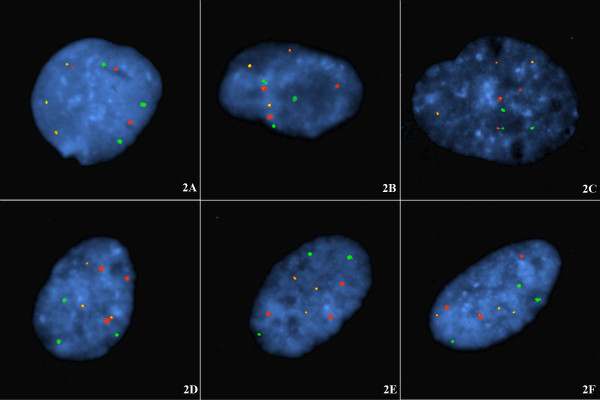
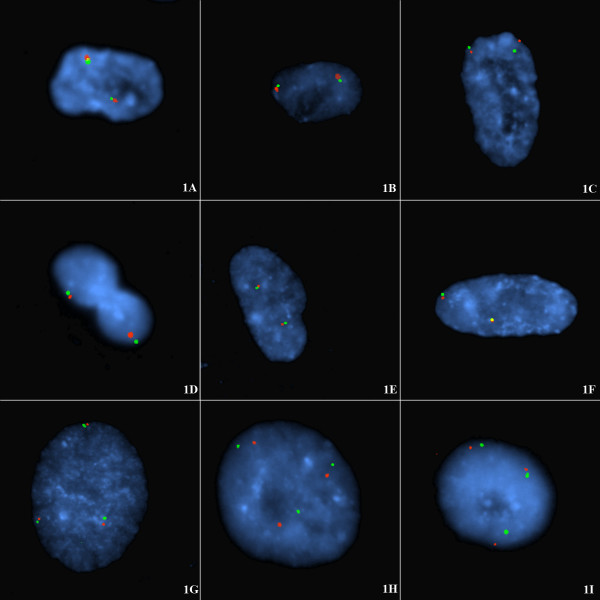
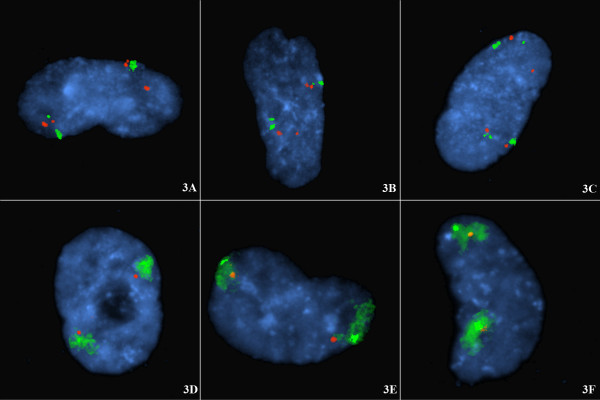
BACKGROUND: To investigate potential mechanisms for telomere capture the spatial arrangement of telomeres and chromosomes was examined in G1 (non-cycling) mitotic cells with diploid or triploid genomes. This was examined firstly by directly labelling the respective short arm (p) and long arm subtelomeres (q) with different fluorophores and probing cell preparations using a number of subtelomere probe pairs, those for chromosomes 1, 3, 4, 5, 6, 7, 9, 10, 12, 17, 18, and 20. In addition some interstitial probes (CEN15, PML and SNRPN on chromosome 15) and whole chromosome paint probes (e.g. WCP12) were jointly hybridised to investigate the co-localization of interphase chromosome domains and tethered subtelomeres. Cells were prepared by omitting exposure to colcemid and hypotonic treatments. RESULTS: In these cells a specific interphase chromosome topology was detected. It was shown that the p and q telomeres of the each chromosome associate frequently (80% pairing) in an intrachromosomal manner, i.e. looped chromosomes with homologues usually widely spaced within the nucleus. This p-q tethering of the telomeres from the one chromosome was observed with large (chromosomes 3, 4, 5), medium sized (6, 7, 9, 10, 12), or small chromosomes (17, 18, 20). When triploid nuclei were probed there were three tetherings of p-q subtelomere signals representing the three widely separated looped chromosome homologues. The separate subtelomere pairings were shown to coincide with separate chromosome domains as defined by the WCP and interstitial probes. The 20% of apparently unpaired subtelomeric signals in diploid nuclei were partially documented to be pairings with the telomeres of other chromosomes. CONCLUSIONS: A topology for telomeres was detected where looped chromosome homologues were present at G1 interphase. These homologues were spatially arranged with respect to one-another independently of other chromosomes, i.e. there was no chromosome order on different sides of the cell nuclei and no segregation into haploid sets was detected. The normal function of this high frequency of intrachromosomal loops is unknown but a potential role is likely in the genesis of telomere captures whether of the intrachromosomal type or between non-homologues. This intrachromosomal tethering of telomeres cannot be related to telomeric or subtelomeric sequences since these are shared in varying degree with other chromosomes. In our view, these intrachromosomal telomeric tetherings with the resulting looped chromosomes arranged in a regular topology must be important to normal cell function since non-cycling cells in G1 are far from quiescent, are in fact metabolically active, and these cells represent the majority status since only a small proportion of cells are normally dividing.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


