Dickson Gerald Mauki, Peter Manning, Matthias Schleuning, Andreas Hemp, David Schellenberger Costa, Natalia Sierra Cornejo, Joscha N. Becker, Andreas Ensslin, Gemma Rutten, Tim Appelhans, Margot Neyret
{"title":"Community-level plant functional strategies explain ecosystem carbon storage across a tropical elevational gradient","authors":"Dickson Gerald Mauki, Peter Manning, Matthias Schleuning, Andreas Hemp, David Schellenberger Costa, Natalia Sierra Cornejo, Joscha N. Becker, Andreas Ensslin, Gemma Rutten, Tim Appelhans, Margot Neyret","doi":"10.1111/1365-2745.70162","DOIUrl":null,"url":null,"abstract":"<h2>1 INTRODUCTION</h2>\n<p>Plant species differ in their allocation of resources to growth, reproduction, and survival, resulting in trait combinations that represent adaptation strategies to various environmental conditions (Chapin III et al., <span>1993</span>; Reich, <span>2014</span>; Wright et al., <span>2004</span>). Global variation in above-ground traits is described in the <i>global spectrum of form and function</i>, which differentiates two principal axes of trait variation. The first contrasts <i>fast</i>, <i>resource-acquisitive</i> strategies with <i>slow</i>, <i>resource-conserving</i> strategies (Wright et al., <span>2004</span>), and the second is related to plant and seed size (Díaz et al., <span>2016</span>; Reich, <span>2014</span>). Additional strategy axes have also been described for below-ground traits, with the <i>collaboration</i> axis describing a gradient of strategies for acquiring soil resources. This ranges from plants with a <i>do-it-yourself</i> strategy that acquire resources via direct root uptake to species <i>outsourcing</i> resource acquisition by forming relationships with mycorrhizal fungi (Bergmann et al., <span>2020</span>; Weigelt et al., <span>2021</span>).</p>\n<p>There is evidence that the major axes of functional strategy variation, initially described for individual species, can also manifest at the community level. This is due to habitat filtering selecting for consistent traits across multiple species of a community (Bruelheide et al., <span>2018</span>). These community-level trait responses are often described using abundance-weighted mean trait values (CWMs). CWMs have been shown to vary in response to environmental gradients across scales. For instance, at a global level, CWMs of plant traits related to a <i>conservative</i> strategy are closely correlated with precipitation gradients, while traits associated with plant size and nutrient acquisition are strongly linked to temperature (Bouchard et al., <span>2024</span>; Joswig et al., <span>2022</span>; Moles et al., <span>2014</span>; Xu et al., <span>2023</span>). Additionally, at the regional scale, Li et al. (<span>2017</span>) observed strong associations between precipitation and CWM root traits related to mycorrhizal colonization, whereas at the local scale, Schellenberger Costa et al. (<span>2017</span>) observed a positive relationship between <i>resource-acquisitive</i> traits and precipitation.</p>\n<p>As plant traits can affect the rate of biogeochemical processes (Chacón-Labella et al., <span>2023</span>), both climate and variation in community-level strategies can influence ecosystem functioning (Freschet et al., <span>2010</span>; Neyret et al., <span>2024</span>; Reich, <span>2014</span>). For instance, Manning et al. (<span>2015</span>) found significant effects of both mean annual temperature and CWM leaf nitrogen content on soil carbon, and Huxley et al. (<span>2023</span>) found significant direct and indirect trait-mediated effects of climate on above-ground net primary productivity. Yet, most of these studies focus only on a few individual traits, often leaf traits, rather than whole-plant strategy variation, and most of these studies do not incorporate root traits (Carmona et al., <span>2021</span>; Cebrián-Piqueras et al., <span>2021</span>; Freschet et al., <span>2010</span>). This is likely due to technical difficulties in root trait measurement and a corresponding lack of data, especially in understudied regions (De Deyn et al., <span>2008</span>; Iversen et al., <span>2017</span>; Laliberté, <span>2017</span>). As a result, there are still major gaps in understanding the relative importance of direct and indirect, trait-mediated climate effects in driving ecosystem functioning, especially the relative roles of both above- and below-ground plant functional traits.</p>\n<p>Here, we focus on one aspect of ecosystem functioning, carbon storage, which plays a crucial role in climate change mitigation (Houghton, <span>2007</span>; Lewis et al., <span>2004</span>). Growing evidence indicates that community-level (CWM) trait variation influences carbon storage (Conti & Díaz, <span>2013</span>; Freschet et al., <span>2012</span>; Vargas-Larreta et al., <span>2021</span>). Specifically, the amount of wood and its carbon content are primary drivers of both above- and below-ground carbon storage (Conti & Díaz, <span>2013</span>). Accordingly, traits linked to biomass production and increased structural investment per unit biomass are expected to directly impact carbon storage (Baker et al., <span>2004</span>; Moles et al., <span>2009</span>). Several studies have reported associations between above-ground CWM traits and carbon, including that between taller vegetation with higher leaf carbon concentrations and higher carbon stocks (Shen, Gilbert, et al., <span>2019</span>; Yang et al., <span>2019</span>). Additionally, root traits can also influence both above-ground and soil carbon storage through their contribution to the <i>slow-fast</i> or <i>collaboration</i> strategy axes. Root traits related to the <i>conservation</i> and <i>outsourcing axes</i> (like high dry matter content and lifespan) have been associated with high carbon storage, while those related to <i>do-it-yourself</i> and <i>fast</i> strategy axis (like specific root length and root nitrogen concentration) have been associated with high-resource turnover and low carbon storage (Lachaise et al., <span>2022</span>; Wen et al., <span>2022</span>). However, while only a few studies have assessed root trait-carbon storage relationships, even fewer have incorporated both above-ground and below-ground traits (but see Chanteloup & Bonis, <span>2013</span>; Craine et al., <span>2002</span>; Orwin et al., <span>2010</span>).</p>\n<p>While many trait–carbon relationships have been described, these are not universally consistent (Feng & Dietze, <span>2013</span>). This may be due to scaling issues, but also because the drivers of carbon storage vary across ecosystem types, with land management and with climatic contexts (Peters et al., <span>2019</span>; Van der Plas et al., <span>2020</span>). In addition to well-known trait responses to climatic conditions, land use can also drive variation in plant functional strategies (Allan et al., <span>2015</span>). Intensive land use, including agricultural and grazing activities, often causes a shift towards species with rapid growth and high reproductive output at the expense of species with <i>conservative</i> traits and longer lifespans (Flynn et al., <span>2011</span>; Laliberte et al., <span>2010</span>; Schellenberger Costa et al., <span>2017</span>). This shift can reduce carbon storage as these fast-growing species typically have lower wood density and shorter root lifespans, leading to faster carbon pool turnover (Garnier et al., <span>2004</span>). Moreover, changes in land use can disrupt relationships between plants and soil symbionts (De Deyn et al., <span>2008</span>), and can alter nutrient availability, soil chemistry, and microbial activity. These soil characteristics may also play an important role in carbon cycling, both directly and by influencing plant community functional trait composition (Joswig et al., <span>2022</span>) and mediating the effects of climate and land use (de Castilho et al., <span>2006</span>; Hartley et al., <span>2021</span>; Manning et al., <span>2015</span>).</p>\n<p>Trait-carbon relationships may also differ between biomes (De Deyn et al., <span>2008</span>). For instance, while above-ground traits may explain carbon storage in above-ground stocks in forests, below-ground traits might better explain carbon stocks in systems where below-ground biomass and resource allocation are high, such as grasslands (Ottaviani et al., <span>2020</span>). This may mean that relationships described in temperate systems cannot be transferred to other biomes, such as tropical ecosystems, which play a major role in carbon storage worldwide. Yet evidence for trait-functioning relationships from these systems is scarce (Finegan et al., <span>2015</span>). Therefore, there is a need for greater understanding of how plant functional strategies interact with land-use intensity and climatic conditions to influence carbon storage in tropical ecosystems.</p>\n<p>Here, we assessed variation in trait composition at the community level and its relationship with above- and below-ground organic carbon storage along the large tropical elevational gradient of Mount Kilimanjaro, which ranges from savanna through forest to alpine ecosystems. This gradient also encompasses variations in land use, as ecosystems at low to mid-elevation have experienced habitat disturbances and land-use transformation (Misana, <span>2012</span>). We hypothesised that coordination of below-ground and above-ground plant traits exists at the community level along the climatic gradients of Kilimanjaro, with communities with <i>acquisitive</i> strategies found in ecosystems with high precipitation, high land-use intensity, and moderate temperature, and communities with <i>conservative</i> strategies found in ecosystems with low precipitation and more extreme temperatures (Hypothesis 1).</p>\n<p>Furthermore, we investigated whether the interplay between climate variables and plant functional strategies was related to patterns of ecosystem carbon storage using structural equation models (Figure 1, where the model structure is also justified). Here we hypothesised that climate influences carbon storage mostly via indirect pathways, mediated by land use and trait effects, with the highest carbon storage being in communities of tall, <i>conservative</i> plants (Hypothesis 2). The hypotheses were tested using vegetation and soil data from 60 plots located along the elevational gradients of Mount Kilimanjaro.</p>\n<figure><picture>\n<source media=\"(min-width: 1650px)\" srcset=\"/cms/asset/af63d1c3-8fd7-457f-8271-2e19b8ce9787/jec70162-fig-0001-m.jpg\"/><img alt=\"Details are in the caption following the image\" data-lg-src=\"/cms/asset/af63d1c3-8fd7-457f-8271-2e19b8ce9787/jec70162-fig-0001-m.jpg\" loading=\"lazy\" src=\"/cms/asset/bc7ad4d1-5365-4234-9359-77293dd36bd1/jec70162-fig-0001-m.png\" title=\"Details are in the caption following the image\"/></picture><figcaption>\n<div><strong>FIGURE 1<span style=\"font-weight:normal\"></span></strong><div>Open in figure viewer<i aria-hidden=\"true\"></i><span>PowerPoint</span></div>\n</div>\n<div>Conceptual model illustrating the plant functional strategies-carbon relationship. Arrows marked I represent direct influences of local climate on carbon storage, arrows marked II represent direct effects of climate on land-use intensity, as on Kilimanjaro, intensive land uses are located at low- to mid-elevation (i.e. warmer plots; Alemayehu et al., <span>2023</span>). Arrow III represents indirect effects of climate on plant functional strategies via land use (Stampfli et al., <span>2018</span>), arrow IV shows the direct of influence of climate on plant strategies (Shen et al., <span>2016</span>), arrow V represents the direct effect of land use on carbon storage (Ensslin et al., <span>2015</span>), and arrow VI represents the influence of functional strategies on carbon storage (Chave et al., <span>2009</span>). Additional models employed a modified version of the structure shown here to investigate the potential role of soil covariates in mediating these relationships (see Supporting Information: Note 3).</div>\n</figcaption>\n</figure>","PeriodicalId":191,"journal":{"name":"Journal of Ecology","volume":"63 1","pages":""},"PeriodicalIF":5.6000,"publicationDate":"2025-09-26","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"Journal of Ecology","FirstCategoryId":"93","ListUrlMain":"https://doi.org/10.1111/1365-2745.70162","RegionNum":1,"RegionCategory":"环境科学与生态学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"ECOLOGY","Score":null,"Total":0}
引用次数: 0
Abstract
1 INTRODUCTION
Plant species differ in their allocation of resources to growth, reproduction, and survival, resulting in trait combinations that represent adaptation strategies to various environmental conditions (Chapin III et al., 1993; Reich, 2014; Wright et al., 2004). Global variation in above-ground traits is described in the global spectrum of form and function, which differentiates two principal axes of trait variation. The first contrasts fast, resource-acquisitive strategies with slow, resource-conserving strategies (Wright et al., 2004), and the second is related to plant and seed size (Díaz et al., 2016; Reich, 2014). Additional strategy axes have also been described for below-ground traits, with the collaboration axis describing a gradient of strategies for acquiring soil resources. This ranges from plants with a do-it-yourself strategy that acquire resources via direct root uptake to species outsourcing resource acquisition by forming relationships with mycorrhizal fungi (Bergmann et al., 2020; Weigelt et al., 2021).
There is evidence that the major axes of functional strategy variation, initially described for individual species, can also manifest at the community level. This is due to habitat filtering selecting for consistent traits across multiple species of a community (Bruelheide et al., 2018). These community-level trait responses are often described using abundance-weighted mean trait values (CWMs). CWMs have been shown to vary in response to environmental gradients across scales. For instance, at a global level, CWMs of plant traits related to a conservative strategy are closely correlated with precipitation gradients, while traits associated with plant size and nutrient acquisition are strongly linked to temperature (Bouchard et al., 2024; Joswig et al., 2022; Moles et al., 2014; Xu et al., 2023). Additionally, at the regional scale, Li et al. (2017) observed strong associations between precipitation and CWM root traits related to mycorrhizal colonization, whereas at the local scale, Schellenberger Costa et al. (2017) observed a positive relationship between resource-acquisitive traits and precipitation.
As plant traits can affect the rate of biogeochemical processes (Chacón-Labella et al., 2023), both climate and variation in community-level strategies can influence ecosystem functioning (Freschet et al., 2010; Neyret et al., 2024; Reich, 2014). For instance, Manning et al. (2015) found significant effects of both mean annual temperature and CWM leaf nitrogen content on soil carbon, and Huxley et al. (2023) found significant direct and indirect trait-mediated effects of climate on above-ground net primary productivity. Yet, most of these studies focus only on a few individual traits, often leaf traits, rather than whole-plant strategy variation, and most of these studies do not incorporate root traits (Carmona et al., 2021; Cebrián-Piqueras et al., 2021; Freschet et al., 2010). This is likely due to technical difficulties in root trait measurement and a corresponding lack of data, especially in understudied regions (De Deyn et al., 2008; Iversen et al., 2017; Laliberté, 2017). As a result, there are still major gaps in understanding the relative importance of direct and indirect, trait-mediated climate effects in driving ecosystem functioning, especially the relative roles of both above- and below-ground plant functional traits.
Here, we focus on one aspect of ecosystem functioning, carbon storage, which plays a crucial role in climate change mitigation (Houghton, 2007; Lewis et al., 2004). Growing evidence indicates that community-level (CWM) trait variation influences carbon storage (Conti & Díaz, 2013; Freschet et al., 2012; Vargas-Larreta et al., 2021). Specifically, the amount of wood and its carbon content are primary drivers of both above- and below-ground carbon storage (Conti & Díaz, 2013). Accordingly, traits linked to biomass production and increased structural investment per unit biomass are expected to directly impact carbon storage (Baker et al., 2004; Moles et al., 2009). Several studies have reported associations between above-ground CWM traits and carbon, including that between taller vegetation with higher leaf carbon concentrations and higher carbon stocks (Shen, Gilbert, et al., 2019; Yang et al., 2019). Additionally, root traits can also influence both above-ground and soil carbon storage through their contribution to the slow-fast or collaboration strategy axes. Root traits related to the conservation and outsourcing axes (like high dry matter content and lifespan) have been associated with high carbon storage, while those related to do-it-yourself and fast strategy axis (like specific root length and root nitrogen concentration) have been associated with high-resource turnover and low carbon storage (Lachaise et al., 2022; Wen et al., 2022). However, while only a few studies have assessed root trait-carbon storage relationships, even fewer have incorporated both above-ground and below-ground traits (but see Chanteloup & Bonis, 2013; Craine et al., 2002; Orwin et al., 2010).
While many trait–carbon relationships have been described, these are not universally consistent (Feng & Dietze, 2013). This may be due to scaling issues, but also because the drivers of carbon storage vary across ecosystem types, with land management and with climatic contexts (Peters et al., 2019; Van der Plas et al., 2020). In addition to well-known trait responses to climatic conditions, land use can also drive variation in plant functional strategies (Allan et al., 2015). Intensive land use, including agricultural and grazing activities, often causes a shift towards species with rapid growth and high reproductive output at the expense of species with conservative traits and longer lifespans (Flynn et al., 2011; Laliberte et al., 2010; Schellenberger Costa et al., 2017). This shift can reduce carbon storage as these fast-growing species typically have lower wood density and shorter root lifespans, leading to faster carbon pool turnover (Garnier et al., 2004). Moreover, changes in land use can disrupt relationships between plants and soil symbionts (De Deyn et al., 2008), and can alter nutrient availability, soil chemistry, and microbial activity. These soil characteristics may also play an important role in carbon cycling, both directly and by influencing plant community functional trait composition (Joswig et al., 2022) and mediating the effects of climate and land use (de Castilho et al., 2006; Hartley et al., 2021; Manning et al., 2015).
Trait-carbon relationships may also differ between biomes (De Deyn et al., 2008). For instance, while above-ground traits may explain carbon storage in above-ground stocks in forests, below-ground traits might better explain carbon stocks in systems where below-ground biomass and resource allocation are high, such as grasslands (Ottaviani et al., 2020). This may mean that relationships described in temperate systems cannot be transferred to other biomes, such as tropical ecosystems, which play a major role in carbon storage worldwide. Yet evidence for trait-functioning relationships from these systems is scarce (Finegan et al., 2015). Therefore, there is a need for greater understanding of how plant functional strategies interact with land-use intensity and climatic conditions to influence carbon storage in tropical ecosystems.
Here, we assessed variation in trait composition at the community level and its relationship with above- and below-ground organic carbon storage along the large tropical elevational gradient of Mount Kilimanjaro, which ranges from savanna through forest to alpine ecosystems. This gradient also encompasses variations in land use, as ecosystems at low to mid-elevation have experienced habitat disturbances and land-use transformation (Misana, 2012). We hypothesised that coordination of below-ground and above-ground plant traits exists at the community level along the climatic gradients of Kilimanjaro, with communities with acquisitive strategies found in ecosystems with high precipitation, high land-use intensity, and moderate temperature, and communities with conservative strategies found in ecosystems with low precipitation and more extreme temperatures (Hypothesis 1).
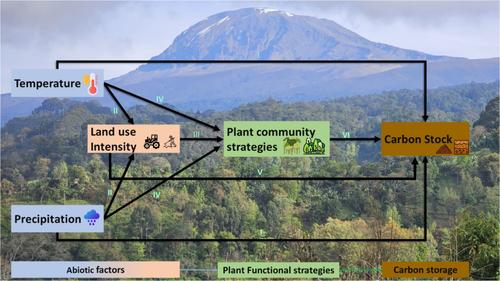
Furthermore, we investigated whether the interplay between climate variables and plant functional strategies was related to patterns of ecosystem carbon storage using structural equation models (Figure 1, where the model structure is also justified). Here we hypothesised that climate influences carbon storage mostly via indirect pathways, mediated by land use and trait effects, with the highest carbon storage being in communities of tall, conservative plants (Hypothesis 2). The hypotheses were tested using vegetation and soil data from 60 plots located along the elevational gradients of Mount Kilimanjaro.
FIGURE 1
Open in figure viewerPowerPoint
Conceptual model illustrating the plant functional strategies-carbon relationship. Arrows marked I represent direct influences of local climate on carbon storage, arrows marked II represent direct effects of climate on land-use intensity, as on Kilimanjaro, intensive land uses are located at low- to mid-elevation (i.e. warmer plots; Alemayehu et al., 2023). Arrow III represents indirect effects of climate on plant functional strategies via land use (Stampfli et al., 2018), arrow IV shows the direct of influence of climate on plant strategies (Shen et al., 2016), arrow V represents the direct effect of land use on carbon storage (Ensslin et al., 2015), and arrow VI represents the influence of functional strategies on carbon storage (Chave et al., 2009). Additional models employed a modified version of the structure shown here to investigate the potential role of soil covariates in mediating these relationships (see Supporting Information: Note 3).
植物物种对生长、繁殖和生存的资源分配不同,导致性状组合代表了对各种环境条件的适应策略(Chapin III et al., 1993; Reich, 2014; Wright et al., 2004)。地上性状的全球变异是用形式和功能的全球谱来描述的,它区分了性状变异的两个主轴。第一个对比了快速的资源获取策略与缓慢的资源节约策略(Wright等人,2004),第二个与植物和种子大小有关(Díaz等人,2016;Reich, 2014)。此外,还描述了地下性状的其他策略轴,其中协作轴描述了获取土壤资源的策略梯度。这包括通过直接根吸收获取资源的自己动手策略的植物,以及通过与菌根真菌形成关系来获取物种外包资源的植物(Bergmann et al., 2020; Weigelt et al., 2021)。有证据表明,最初描述为单个物种的功能策略变化的主要轴也可以在群落水平上表现出来。这是由于栖息地过滤选择了一个群落中多个物种的一致特征(Bruelheide et al., 2018)。这些社区水平的性状反应通常用丰度加权平均性状值(CWMs)来描述。CWMs已被证明对不同尺度的环境梯度有不同的响应。例如,在全球水平上,与保守策略相关的植物性状的CWMs与降水梯度密切相关,而与植物大小和养分获取相关的性状与温度密切相关(Bouchard et al., 2024; Joswig et al., 2022; mole et al., 2014; Xu et al., 2023)。此外,在区域尺度上,Li等人(2017)观察到降水与与菌根定植相关的CWM根系性状之间存在很强的相关性,而在局部尺度上,Schellenberger Costa等人(2017)观察到资源获取性状与降水之间存在正相关关系。由于植物性状可以影响生物地球化学过程的速率(Chacón-Labella等人,2023),气候和群落层面策略的变化都可以影响生态系统功能(Freschet等人,2010;Neyret等人,2024;Reich, 2014)。例如,Manning等人(2015)发现年平均温度和CWM叶片氮含量对土壤碳都有显著影响,Huxley等人(2023)发现气候对地上净初级生产力有显著的直接和间接性状介导的影响。然而,这些研究大多只关注少数个体性状,通常是叶片性状,而不是整个植物的策略变异,而且大多数研究没有纳入根性状(Carmona et al., 2021; Cebrián-Piqueras et al., 2021; Freschet et al., 2010)。这可能是由于根性状测量的技术困难和相应的数据缺乏,特别是在研究不足的地区(De Deyn等人,2008;Iversen等人,2017;lalibert<e:1>, 2017)。因此,在了解直接和间接、性状介导的气候效应在驱动生态系统功能中的相对重要性,特别是地上和地下植物功能性状的相对作用方面仍然存在重大差距。在这里,我们重点关注生态系统功能的一个方面,即碳储存,它在减缓气候变化方面起着至关重要的作用(Houghton, 2007; Lewis等人,2004)。越来越多的证据表明,社区水平(CWM)性状变异会影响碳储量(Conti & Díaz, 2013; Freschet et al., 2012; varga - larreta et al., 2021)。具体来说,木材的数量及其碳含量是地上和地下碳储存的主要驱动因素(Conti & Díaz, 2013)。因此,与生物量生产相关的特征和单位生物量结构投资的增加预计将直接影响碳储量(Baker等人,2004;mole等人,2009)。一些研究报告了地上CWM性状与碳之间的关联,包括叶片碳浓度较高的较高植被与较高碳储量之间的关联(Shen, Gilbert, et al., 2019; Yang et al., 2019)。此外,根系性状还可以通过对慢速策略轴或协同策略轴的贡献来影响地上和土壤碳储量。与保护和外包轴相关的根系性状(如高干物质含量和寿命)与高碳储量相关,而与自己动手和快速策略轴相关的根系性状(如比根长度和根氮浓度)与高资源周转率和低碳储量相关(Lachaise等,2022;Wen等,2022)。 然而,虽然只有少数研究评估了根系性状与碳储量的关系,但将地上和地下性状结合起来的研究就更少了(但参见Chanteloup &; Bonis, 2013; Craine et al., 2002; Orwin et al., 2010)。虽然已经描述了许多性状与碳的关系,但这些关系并不普遍一致(Feng & Dietze, 2013)。这可能是由于尺度问题,但也因为碳储存的驱动因素因生态系统类型、土地管理和气候背景而异(Peters等人,2019;Van der Plas等人,2020)。除了众所周知的性状对气候条件的响应外,土地利用还可以驱动植物功能策略的变化(Allan et al., 2015)。集约土地利用,包括农业和放牧活动,往往导致物种向生长迅速、繁殖产量高的物种转移,而牺牲了性状保守、寿命较长的物种(Flynn等人,2011;Laliberte等人,2010;Schellenberger Costa等人,2017)。这种转变可以减少碳储量,因为这些快速生长的物种通常具有较低的木材密度和较短的根系寿命,从而导致更快的碳库周转(Garnier et al., 2004)。此外,土地利用的变化会破坏植物与土壤共生体之间的关系(De Deyn et al., 2008),并会改变养分有效性、土壤化学和微生物活动。这些土壤特征也可能在碳循环中发挥重要作用,直接或通过影响植物群落功能性状组成(Joswig等人,2022)和调节气候和土地利用的影响(de Castilho等人,2006;Hartley等人,2021;Manning等人,2015)。性状-碳关系在不同的生物群落之间也可能不同(De Deyn et al., 2008)。例如,虽然地上性状可以解释森林中地上储量的碳储量,但地下性状可能更好地解释地下生物量和资源分配高的系统(如草地)中的碳储量(Ottaviani et al., 2020)。这可能意味着在温带系统中描述的关系不能转移到其他生物群系,例如在全球碳储存中起主要作用的热带生态系统。然而,这些系统中特征-功能关系的证据很少(Finegan et al., 2015)。因此,有必要进一步了解植物功能策略如何与土地利用强度和气候条件相互作用,从而影响热带生态系统的碳储量。本研究在乞力马扎罗山沿热带大海拔梯度,从热带稀树草原到森林再到高山生态系统,评估了群落特征组成的变化及其与地上和地下有机碳储量的关系。这种梯度还包括土地利用的变化,因为中低海拔的生态系统经历了栖息地的干扰和土地利用的转变(Misana, 2012)。我们假设,在乞力马扎罗山的气候梯度上,植物地下和地上性状在群落水平上存在协调,在高降水、高土地利用强度和中等温度的生态系统中,群落具有获取策略,而在低降水和更极端温度的生态系统中,群落具有保守策略(假设1)。此外,我们利用结构方程模型研究了气候变量与植物功能策略之间的相互作用是否与生态系统碳储量模式有关(图1,其中模型结构也得到了证明)。在这里,我们假设气候主要通过间接途径影响碳储量,由土地利用和性状效应介导,在高大、保守的植物群落中碳储量最高(假设2)。利用乞力马扎罗山海拔梯度沿线60个样地的植被和土壤数据对这些假设进行了检验。图1打开图形查看器powerpoint说明植物功能策略的概念模型-碳关系。标记为I的箭头表示当地气候对碳储量的直接影响,标记为II的箭头表示气候对土地利用强度的直接影响,因为在乞力马扎罗山,集约土地利用位于中低海拔(即较温暖的地块;Alemayehu等人,2023)。箭头III表示气候通过土地利用间接影响植物的功能策略(Stampfli et al., 2018),箭头IV表示气候对植物策略的直接影响(Shen et al., 2016),箭头V表示土地利用对碳储量的直接影响(Ensslin et al., 2015),箭头VI表示功能策略对碳储量的影响(Chave et al., 2009)。 其他模型采用此处所示结构的修改版本来研究土壤协变量在中介这些关系中的潜在作用(见支持信息:注3)。
期刊介绍:
Journal of Ecology publishes original research papers on all aspects of the ecology of plants (including algae), in both aquatic and terrestrial ecosystems. We do not publish papers concerned solely with cultivated plants and agricultural ecosystems. Studies of plant communities, populations or individual species are accepted, as well as studies of the interactions between plants and animals, fungi or bacteria, providing they focus on the ecology of the plants.
We aim to bring important work using any ecological approach (including molecular techniques) to a wide international audience and therefore only publish papers with strong and ecological messages that advance our understanding of ecological principles.


 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


