{"title":"The SLC58A Na+-Monocarboxylate Transporter—Can It Scavenge Lung Metabolites to Prevent Airway Infections?","authors":"Deborah Baines, Robert Tarran","doi":"10.1111/apha.70086","DOIUrl":null,"url":null,"abstract":"<p>The airway surface liquid (ASL), which lines the luminal surface of the lung, is a complex layer containing mucins that trap inhaled particles and a liquid layer that supports ciliary function, that also contains antimicrobial peptides, proteins, and metabolites generated by the epithelial cells, inflammatory cells, and the resident lung microbiota. Precise regulation of ASL composition protects the pulmonary tissue from the external environment and is critical for a healthy lung.</p><p>Much is known about the identity and function of the airway epithelial ion channels and transporters that contribute to the regulation of ASL volume and mucus clearance. Nucleotides and nucleosides in the ASL modify fluid volume through receptor-mediatedion transport mechanisms [<span>1</span>], while bacterial metabolites are sensed by taste receptors in the ciliated and chemosensory cells of the airway and initiate protective reflexes [<span>2</span>].</p><p>Intermediary metabolites of glucose, such as lactate and pyruvate, as well as short chain fatty acids, are often elevated in the ASL during disease, and can render the lung more susceptible to infection and/or inflammation [<span>3</span>]. The increased abundance of ASL metabolites is associated with changes in cellular synthesis and transport [<span>4</span>]. The production and secretion of L-lactate into the ASL increase during hyperglycaemia, in the presence of bacteria and inflammation [<span>5, 6</span>]. The role of the H<sup>+</sup>-coupled monocarboxylate transporters (e.g., SLC16A1, 7 and 3; MCT1, 2 and 4 respectively) in the secretion of such metabolites, including into the ASL, has been reported [<span>5, 7</span>]. But is this a one-way process? Are metabolites also removed from the ASL and if so, how? A recent editorial and manuscript in Acta Physiologica highlighted new views on the shuttling of lactate from cell to cell and tissue to tissue as a proposed energy source, supporting both its secretion and uptake [<span>8, 9</span>]. But until now, there has been little documented evidence for transporters that enable the uptake of metabolites across the lumen of the airway and could play a role in the regulation of ASL metabolite concentration.</p><p>A new manuscript in Acta Physiologica, by Guenquen et al. [<span>10</span>] has changed that. The authors found that the Na<sup>+</sup>-coupled monocarboxylate transporter SLC5A8 (SMCT1) was highly expressed in mouse trachea and in human bronchial epithelial cultures. SLC5A8 is a member of a family that also includes more well-known Na<sup>+</sup>-coupled glucose transporters, such as SLC5A1 (SGLT1). These transporters can utilize the Na<sup>+</sup> gradient established by the epithelial Na<sup>+</sup>/K<sup>+</sup>-ATPase to drive uptake into the cell in the absence of a substrate gradient. As highlighted by Guenquen and colleagues, SLC5A8 has previously attracted attention as a tumor suppressor in several tissues (including the lung) and has been documented to transport lactate and butyrate in the kidney and colon, respectively.</p><p>The authors elegantly show that along with Na<sup>+</sup>, SLC5A8 electrogenically transports a broad range of metabolites including D- and L-lactate, pyruvate, acetate, propionate, and nicotinate across the luminal epithelial membrane. They also provide evidence that SLC58A is expressed in key epithelial cells alongside channels such as the epithelial Na<sup>+</sup> channel (ENaC) and the cystic fibrosis transmembrane conductance regulator (CFTR), which regulate ASL volume.</p><p>The authors offer new evidence that SLC5A8 plays a key role in regulating L-lactate and other metabolite concentrations in the ASL. The authors show that SLC5A8 exhibited transport activity over a wide range of metabolite concentrations and that SLC5A8 expression was increased in response to pro-inflammatory stimuli. Taken together, these data suggest that SLC5A8 transports metabolites such as L-lactate at relevant concentrations found in the ASL, and that SLC5A8 upregulation in response to inflammation could increase monocarboxylate uptake to mitigate the elevated metabolite concentrations generated by increased cellular metabolism during infection. Thus, SLC5A8 provides a new mechanism for the control of ASL metabolite concentrations (Figure 1). Further extrapolation suggests that since increased SLC5A8 activity lowers ASL metabolite concentrations, SLC5A8 has the potential to modify the inflammatory response and contribute to the maintenance of the innate immune function of the airway.</p><p>As the authors allude, this discovery has opened a cornucopia of new questions. The co-transport of Na<sup>+</sup> by SLC5A8 (like SGLT1) is likely to modify ASL volume absorption (Figure 1), which is critical for the maintenance of the mucociliary escalator. The authors showed that electrogenic Na<sup>+</sup> transport was not additive to the function of ENaC, the main driver of fluid absorption. But, as they suggest, it remains to be seen how SLC5A8 activity interacts with other channels, transporters, and signaling pathways to regulate ASL hydration in normal and disease conditions. Interestingly, SLC5A1 was recently shown to be upregulated in cystic fibrosis (CF) airways, modifying airway hydration and, as such, is being proposed as a potential therapeutic target [<span>11</span>].</p><p>The differential response to inhibition with valproate indicates that human and murine SLC5A8 may differ in structure with regulatory and physiological consequences that need to be understood. How human SLC5A8 is regulated, and how it coordinates with other monocarboxylate transporters to scavenge metabolites from the ASL to moderate the effects of inflammation/infection need to be unraveled. How SLC5A8 functions in airway disease (e.g., asthma, chronic obstructive pulmonary disease, CF) where both metabolic changes and inflammation are evident, is an exciting new avenue for exploration. Perhaps a key question for the future is whether SLC5A8 could be pharmacologically manipulated to mitigate metabolite-linked inflammatory responses. Watch this space!</p><p><b>Deborah Baines:</b> writing – original draft. <b>Robert Tarran:</b> writing – review and editing.</p><p>The authors declare no conflicts of interest.</p>","PeriodicalId":107,"journal":{"name":"Acta Physiologica","volume":"241 9","pages":""},"PeriodicalIF":5.6000,"publicationDate":"2025-08-02","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1111/apha.70086","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"Acta Physiologica","FirstCategoryId":"3","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1111/apha.70086","RegionNum":2,"RegionCategory":"医学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"PHYSIOLOGY","Score":null,"Total":0}
引用次数: 0
Abstract
The airway surface liquid (ASL), which lines the luminal surface of the lung, is a complex layer containing mucins that trap inhaled particles and a liquid layer that supports ciliary function, that also contains antimicrobial peptides, proteins, and metabolites generated by the epithelial cells, inflammatory cells, and the resident lung microbiota. Precise regulation of ASL composition protects the pulmonary tissue from the external environment and is critical for a healthy lung.
Much is known about the identity and function of the airway epithelial ion channels and transporters that contribute to the regulation of ASL volume and mucus clearance. Nucleotides and nucleosides in the ASL modify fluid volume through receptor-mediatedion transport mechanisms [1], while bacterial metabolites are sensed by taste receptors in the ciliated and chemosensory cells of the airway and initiate protective reflexes [2].
Intermediary metabolites of glucose, such as lactate and pyruvate, as well as short chain fatty acids, are often elevated in the ASL during disease, and can render the lung more susceptible to infection and/or inflammation [3]. The increased abundance of ASL metabolites is associated with changes in cellular synthesis and transport [4]. The production and secretion of L-lactate into the ASL increase during hyperglycaemia, in the presence of bacteria and inflammation [5, 6]. The role of the H+-coupled monocarboxylate transporters (e.g., SLC16A1, 7 and 3; MCT1, 2 and 4 respectively) in the secretion of such metabolites, including into the ASL, has been reported [5, 7]. But is this a one-way process? Are metabolites also removed from the ASL and if so, how? A recent editorial and manuscript in Acta Physiologica highlighted new views on the shuttling of lactate from cell to cell and tissue to tissue as a proposed energy source, supporting both its secretion and uptake [8, 9]. But until now, there has been little documented evidence for transporters that enable the uptake of metabolites across the lumen of the airway and could play a role in the regulation of ASL metabolite concentration.
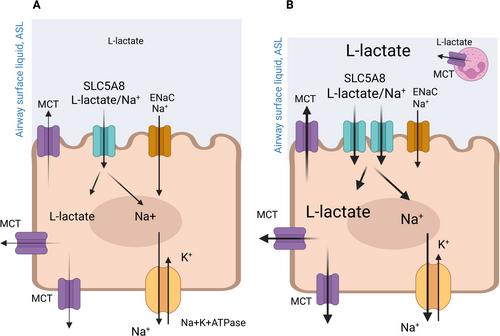
A new manuscript in Acta Physiologica, by Guenquen et al. [10] has changed that. The authors found that the Na+-coupled monocarboxylate transporter SLC5A8 (SMCT1) was highly expressed in mouse trachea and in human bronchial epithelial cultures. SLC5A8 is a member of a family that also includes more well-known Na+-coupled glucose transporters, such as SLC5A1 (SGLT1). These transporters can utilize the Na+ gradient established by the epithelial Na+/K+-ATPase to drive uptake into the cell in the absence of a substrate gradient. As highlighted by Guenquen and colleagues, SLC5A8 has previously attracted attention as a tumor suppressor in several tissues (including the lung) and has been documented to transport lactate and butyrate in the kidney and colon, respectively.
The authors elegantly show that along with Na+, SLC5A8 electrogenically transports a broad range of metabolites including D- and L-lactate, pyruvate, acetate, propionate, and nicotinate across the luminal epithelial membrane. They also provide evidence that SLC58A is expressed in key epithelial cells alongside channels such as the epithelial Na+ channel (ENaC) and the cystic fibrosis transmembrane conductance regulator (CFTR), which regulate ASL volume.
The authors offer new evidence that SLC5A8 plays a key role in regulating L-lactate and other metabolite concentrations in the ASL. The authors show that SLC5A8 exhibited transport activity over a wide range of metabolite concentrations and that SLC5A8 expression was increased in response to pro-inflammatory stimuli. Taken together, these data suggest that SLC5A8 transports metabolites such as L-lactate at relevant concentrations found in the ASL, and that SLC5A8 upregulation in response to inflammation could increase monocarboxylate uptake to mitigate the elevated metabolite concentrations generated by increased cellular metabolism during infection. Thus, SLC5A8 provides a new mechanism for the control of ASL metabolite concentrations (Figure 1). Further extrapolation suggests that since increased SLC5A8 activity lowers ASL metabolite concentrations, SLC5A8 has the potential to modify the inflammatory response and contribute to the maintenance of the innate immune function of the airway.
As the authors allude, this discovery has opened a cornucopia of new questions. The co-transport of Na+ by SLC5A8 (like SGLT1) is likely to modify ASL volume absorption (Figure 1), which is critical for the maintenance of the mucociliary escalator. The authors showed that electrogenic Na+ transport was not additive to the function of ENaC, the main driver of fluid absorption. But, as they suggest, it remains to be seen how SLC5A8 activity interacts with other channels, transporters, and signaling pathways to regulate ASL hydration in normal and disease conditions. Interestingly, SLC5A1 was recently shown to be upregulated in cystic fibrosis (CF) airways, modifying airway hydration and, as such, is being proposed as a potential therapeutic target [11].
The differential response to inhibition with valproate indicates that human and murine SLC5A8 may differ in structure with regulatory and physiological consequences that need to be understood. How human SLC5A8 is regulated, and how it coordinates with other monocarboxylate transporters to scavenge metabolites from the ASL to moderate the effects of inflammation/infection need to be unraveled. How SLC5A8 functions in airway disease (e.g., asthma, chronic obstructive pulmonary disease, CF) where both metabolic changes and inflammation are evident, is an exciting new avenue for exploration. Perhaps a key question for the future is whether SLC5A8 could be pharmacologically manipulated to mitigate metabolite-linked inflammatory responses. Watch this space!
Deborah Baines: writing – original draft. Robert Tarran: writing – review and editing.
期刊介绍:
Acta Physiologica is an important forum for the publication of high quality original research in physiology and related areas by authors from all over the world. Acta Physiologica is a leading journal in human/translational physiology while promoting all aspects of the science of physiology. The journal publishes full length original articles on important new observations as well as reviews and commentaries.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


