ARC3 required for constriction
IF 13.6
1区 生物学
Q1 PLANT SCIENCES
引用次数: 0
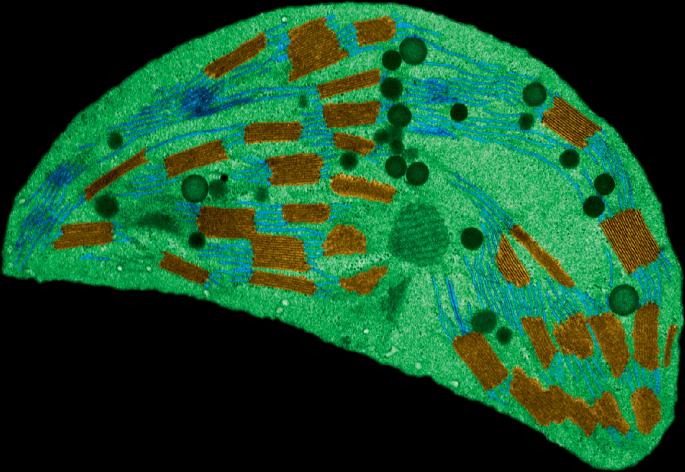
收缩需要ARC3
叶绿体分裂需要形成一个由两个类似微管蛋白的FtsZ同源物FtsZ1和FtsZ2组成的基质Z环。该环通过ARC6连接到内包膜,并通过PLASTID DIVISION蛋白进一步连接到外包膜,并连接到含有动力蛋白样ARC5的细胞质质体分裂环。Z环的正确定位取决于Min系统的叶绿体版本,其中ARC3作为Z环形成的负调节因子起着关键作用。ARC3在非分裂位点显示弥漫性定位,这可能阻止了Z环的形成。然而,ARC3也定位于Z环本身,但该亚群的功能尚不清楚。研究人员发现,ARC3与ARC6存在物理相互作用,这种相互作用依赖于其膜占领和识别连接域(MORN),已知它与ARC6的旁链结合(parac6)。事实上,ARC3在arc6、arc6和arc6突变背景中的定位表明,arc6和arc6在将ARC3招募到Z环方面存在一定的功能重叠。在异源酵母系统中,缺失MORN结构域的ARC3表达可消除FtsZ1的丝组装,这为该结构域的自抑制功能提供了证据。ARC6与ARC3的共表达抑制了FtsZ1纤维的形成,可能与MORN结构域相互作用,从而释放类似于PARC6的自抑制功能。然而,与PARC6相比,ARC6在植物中有一个类似j的结构域,阻止了与ARC3的相互作用。事实上,缺失ARC6的J-like结构域导致FtsZ1组装减少和叶绿体分裂缺陷,类似于过表达ARC3的植物。但在arc3突变体背景下,情况并非如此,这表明FtsZ1组装的抑制依赖于arc3。有趣的是,在异源毕赤酵母系统中,ARC3和ARC6的共表达导致Z环收缩动力学增强,这表明ARC3的功能不仅受到Z环内ARC6的负向控制,而且有利于收缩动力学。
本文章由计算机程序翻译,如有差异,请以英文原文为准。
求助全文
约1分钟内获得全文
求助全文
来源期刊
Nature Plants
PLANT SCIENCES-
CiteScore
25.30
自引率
2.20%
发文量
196
期刊介绍:
Nature Plants is an online-only, monthly journal publishing the best research on plants — from their evolution, development, metabolism and environmental interactions to their societal significance.
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


