{"title":"Lactate Refurbished: Cardiovascular Support During Metabolic Stress and Fuel Rather Than Waste","authors":"Cor de Wit","doi":"10.1111/apha.70064","DOIUrl":null,"url":null,"abstract":"<p>For decades, lactate was considered a metabolic waste product that is produced and released from cells during (relative) lack of oxygen. In the glycolytic pathway, glucose is metabolized to pyruvate that fuels the Krebs cycle in mitochondria where oxygen is required for complete pyruvate oxidation. It was thought that pyruvate accumulates during oxygen lack and is subsequently reduced to lactate with concomitant NADH consumption and regeneration of NAD<sup>+</sup>. The equilibrium of the reaction is determined by the lactate/pyruvate ratio and the cytosolic redox potential (NADH/NAD<sup>+</sup> ratio) leading to 10-fold higher concentrations of lactate than pyruvate in resting skeletal muscle. This generated the idea that lactate is the final product of the glycolytic pathway and, in fact, links glycolysis to oxidative metabolism [<span>1</span>]. Interestingly, lactate is also produced under aerobic conditions, that is at pO<sub>2</sub> values well above the critical mitochondrial pO<sub>2</sub>. Lactate production is even enhanced in exercising skeletal muscle because glycolysis is stimulated more intensely than oxidative phosphorylation [<span>2</span>]. Therefore, lactate is also released into the bloodstream from working muscle providing a dynamic fuel source that may be taken up and metabolized by other organs including brain and heart, which are known to eagerly combust lactate from blood [<span>3</span>].</p><p>This concept turns the spotlight on lactate as an important player in metabolism during exercise. It may not only be shuttled from the cytosol to mitochondria but also from cell to cell and tissue to tissue via the bloodstream [<span>1</span>] providing a valuable energy source for tissues that do not exhibit large energy stores. In this context, the work of Homilius and colleagues published in this issue [<span>4</span>] provides interesting new insight by characterizing the impact of lactate on cardiovascular homeostasis. They demonstrate that adjustments required in the cardiovascular system with enhanced cellular metabolism during exercise are uniquely induced by this valuable metabolite (Figure 1).</p><p>Homilius and colleagues [<span>4</span>] studied the vascular effects of different conjugate bases that are produced during cellular metabolism or by the microbiome. Effects of protons were eliminated by keeping pH constant. Precontracted arteries isolated from different vascular beds from rats relaxed largely or even fully in response to bases of possible microbial origin (acetate, propionate, butyrate) at seemingly high concentrations (10 mmol/L). Endogenously produced bases were less efficacious (lactate, hydroxybutyrate), but still achieved considerable relaxations of 25%–50% in different arteries, with pronounced relaxations in femoral and coronary arteries, after 5 min at concentrations found during exercise (EC50: ~10 mmol/L). Other bases were without substantial effect (pyruvate, malonate, succinate). In marked contrast to most bases, that relaxed arteries and veins, lactate produced mostly constrictions in different rat veins. A similar divergent effect was confirmed in human vessels, that is, relaxation in arteries and constriction in veins. Finally, hemodynamic measurements including echocardiography were obtained in rats. Lactate infusion enhanced plasma levels moderately (to 3.7 mmol/L) and augmented cardiac output by increasing stroke volume (by ~20%). The concomitant rise in end-diastolic volume led the authors conclude that the elevated cardiac performance is primarily due to the lactate induced venous constriction. It discharges blood from veins, promotes cardiac filling and thereby modulates length-dependent cardiac muscle activation (Frank-Starling). This may be supported by a separate positive inotropic effect revealed in isolated rat hearts at constant preload. Notably, calculated systemic vascular resistance decreased during lactate infusion as reflected by unchanged arterial pressure in the face of enhanced cardiac output. Taken together, the elaborate and elegant experiments of Homilius and colleagues suggest that lactate, as they so rightly stated in their title, ‘orchestrates cardiovascular adaptations’ during metabolic challenges such as exercise. This role becomes even more interesting for physiologists if one considers lactate not being waste but rather being fuel. It is released from cells that generate lactate in larger quantities than actually combusted in their mitochondria. In this context, lactate not only provides a fuel but also supports the cardiovascular system to meet the challenges in the face of exercise.</p><p>Does lactate also participate in the dilation in response to exercise (active hyperemia) in the respective tissue itself? The current data do not provide answers to this question. Homilius and colleagues demonstrated that lactate decreased vascular resistance, suggesting that it indeed relaxes resistance vessels and not only those larger arteries studied in vitro. It still seems unlikely that lactate released by exercising skeletal muscle reaches upstream arterioles. If lactate would induce a hyperpolarization in endothelial cells, it may also induce dilations at upstream sites (so-called ascending dilation) [<span>5</span>]. However, currently only little is known about the mechanisms of lactate-induced dilation. Some studies suggest that endothelial NO release is invoked; others suggest Ca<sup>2+</sup>-activated K<sup>+</sup>-channels or increases in cAMP in smooth muscle cells as mediators [<span>6</span>]. These mechanisms are rather unlikely to elicit an ascending dilation. In addition, Homilius and colleagues demonstrated that the interval between application and dilation is rather long (minutes). However, active hyperemia sets in within seconds and reaches maximal levels within less than a minute during skeletal muscle exercise [<span>7</span>]. Also, interstitial lactate concentrations in skeletal muscle increased during exercise in humans and continued to rise during recovery when enhanced blood flow already declined [<span>8</span>]. Thus, other mechanisms are more likely to contribute to this important physiologic response [<span>9</span>].</p><p>The most puzzling aspect is the divergent action of lactate on arteries and veins, which is functionally useful in exercising conditions. Which mechanisms may underlie such a distinct response? This question was not (yet) addressed by the authors. One may speculate that endothelium-derived dilator mechanisms are involved in arteries while venous endothelium lacks respective receptors or transporters, and it is just smooth muscle that is activated, resulting in constriction.</p><p>Finally, lactate is a reliable predictor of poor prognosis in severe pathologic conditions. Hyperlactatemia is commonly attributed in the clinical arena to enhanced release from tissues supposedly being hypoxic subsequent to poor tissue perfusion, while the causing tissues often remain unidentified. However, enhanced release may well just be due to metabolic stress with glycolytic production exceeding lactate drainage to mitochondria. This may be aggravated by enhanced catecholamine levels in critically ill patients [<span>10</span>]. Additionally, lactate uptake and oxidation in other organs may be reduced. Nevertheless, hyperlactatemia remains a predictor for mortality; however, the underlying mechanisms are possibly misunderstood. Therefore, it seems to be worth the effort to reframe the image of lactate in the mind of physiologists and clinicians. Instead of regarding lactate as a waste product, its importance as a metabolite exerting even beneficial effects in the cardiovascular system should be considered, as shown by Homilius and colleagues [<span>4</span>].</p><p><b>Cor de Wit:</b> conceptualization, writing – original draft, writing – review and editing, visualization, funding acquisition.</p><p>The author declares no conflicts of interest.</p>","PeriodicalId":107,"journal":{"name":"Acta Physiologica","volume":"241 7","pages":""},"PeriodicalIF":5.6000,"publicationDate":"2025-05-28","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1111/apha.70064","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"Acta Physiologica","FirstCategoryId":"3","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1111/apha.70064","RegionNum":2,"RegionCategory":"医学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"PHYSIOLOGY","Score":null,"Total":0}
引用次数: 0
Abstract
For decades, lactate was considered a metabolic waste product that is produced and released from cells during (relative) lack of oxygen. In the glycolytic pathway, glucose is metabolized to pyruvate that fuels the Krebs cycle in mitochondria where oxygen is required for complete pyruvate oxidation. It was thought that pyruvate accumulates during oxygen lack and is subsequently reduced to lactate with concomitant NADH consumption and regeneration of NAD+. The equilibrium of the reaction is determined by the lactate/pyruvate ratio and the cytosolic redox potential (NADH/NAD+ ratio) leading to 10-fold higher concentrations of lactate than pyruvate in resting skeletal muscle. This generated the idea that lactate is the final product of the glycolytic pathway and, in fact, links glycolysis to oxidative metabolism [1]. Interestingly, lactate is also produced under aerobic conditions, that is at pO2 values well above the critical mitochondrial pO2. Lactate production is even enhanced in exercising skeletal muscle because glycolysis is stimulated more intensely than oxidative phosphorylation [2]. Therefore, lactate is also released into the bloodstream from working muscle providing a dynamic fuel source that may be taken up and metabolized by other organs including brain and heart, which are known to eagerly combust lactate from blood [3].
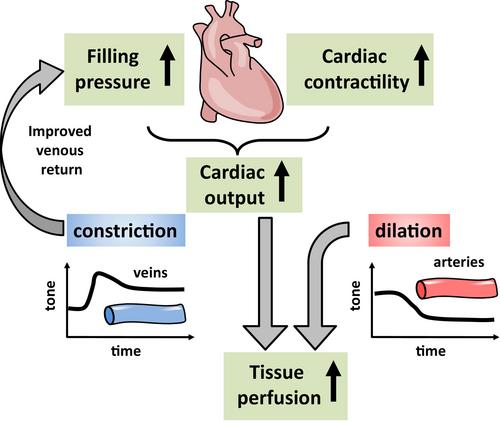
This concept turns the spotlight on lactate as an important player in metabolism during exercise. It may not only be shuttled from the cytosol to mitochondria but also from cell to cell and tissue to tissue via the bloodstream [1] providing a valuable energy source for tissues that do not exhibit large energy stores. In this context, the work of Homilius and colleagues published in this issue [4] provides interesting new insight by characterizing the impact of lactate on cardiovascular homeostasis. They demonstrate that adjustments required in the cardiovascular system with enhanced cellular metabolism during exercise are uniquely induced by this valuable metabolite (Figure 1).
Homilius and colleagues [4] studied the vascular effects of different conjugate bases that are produced during cellular metabolism or by the microbiome. Effects of protons were eliminated by keeping pH constant. Precontracted arteries isolated from different vascular beds from rats relaxed largely or even fully in response to bases of possible microbial origin (acetate, propionate, butyrate) at seemingly high concentrations (10 mmol/L). Endogenously produced bases were less efficacious (lactate, hydroxybutyrate), but still achieved considerable relaxations of 25%–50% in different arteries, with pronounced relaxations in femoral and coronary arteries, after 5 min at concentrations found during exercise (EC50: ~10 mmol/L). Other bases were without substantial effect (pyruvate, malonate, succinate). In marked contrast to most bases, that relaxed arteries and veins, lactate produced mostly constrictions in different rat veins. A similar divergent effect was confirmed in human vessels, that is, relaxation in arteries and constriction in veins. Finally, hemodynamic measurements including echocardiography were obtained in rats. Lactate infusion enhanced plasma levels moderately (to 3.7 mmol/L) and augmented cardiac output by increasing stroke volume (by ~20%). The concomitant rise in end-diastolic volume led the authors conclude that the elevated cardiac performance is primarily due to the lactate induced venous constriction. It discharges blood from veins, promotes cardiac filling and thereby modulates length-dependent cardiac muscle activation (Frank-Starling). This may be supported by a separate positive inotropic effect revealed in isolated rat hearts at constant preload. Notably, calculated systemic vascular resistance decreased during lactate infusion as reflected by unchanged arterial pressure in the face of enhanced cardiac output. Taken together, the elaborate and elegant experiments of Homilius and colleagues suggest that lactate, as they so rightly stated in their title, ‘orchestrates cardiovascular adaptations’ during metabolic challenges such as exercise. This role becomes even more interesting for physiologists if one considers lactate not being waste but rather being fuel. It is released from cells that generate lactate in larger quantities than actually combusted in their mitochondria. In this context, lactate not only provides a fuel but also supports the cardiovascular system to meet the challenges in the face of exercise.
Does lactate also participate in the dilation in response to exercise (active hyperemia) in the respective tissue itself? The current data do not provide answers to this question. Homilius and colleagues demonstrated that lactate decreased vascular resistance, suggesting that it indeed relaxes resistance vessels and not only those larger arteries studied in vitro. It still seems unlikely that lactate released by exercising skeletal muscle reaches upstream arterioles. If lactate would induce a hyperpolarization in endothelial cells, it may also induce dilations at upstream sites (so-called ascending dilation) [5]. However, currently only little is known about the mechanisms of lactate-induced dilation. Some studies suggest that endothelial NO release is invoked; others suggest Ca2+-activated K+-channels or increases in cAMP in smooth muscle cells as mediators [6]. These mechanisms are rather unlikely to elicit an ascending dilation. In addition, Homilius and colleagues demonstrated that the interval between application and dilation is rather long (minutes). However, active hyperemia sets in within seconds and reaches maximal levels within less than a minute during skeletal muscle exercise [7]. Also, interstitial lactate concentrations in skeletal muscle increased during exercise in humans and continued to rise during recovery when enhanced blood flow already declined [8]. Thus, other mechanisms are more likely to contribute to this important physiologic response [9].
The most puzzling aspect is the divergent action of lactate on arteries and veins, which is functionally useful in exercising conditions. Which mechanisms may underlie such a distinct response? This question was not (yet) addressed by the authors. One may speculate that endothelium-derived dilator mechanisms are involved in arteries while venous endothelium lacks respective receptors or transporters, and it is just smooth muscle that is activated, resulting in constriction.
Finally, lactate is a reliable predictor of poor prognosis in severe pathologic conditions. Hyperlactatemia is commonly attributed in the clinical arena to enhanced release from tissues supposedly being hypoxic subsequent to poor tissue perfusion, while the causing tissues often remain unidentified. However, enhanced release may well just be due to metabolic stress with glycolytic production exceeding lactate drainage to mitochondria. This may be aggravated by enhanced catecholamine levels in critically ill patients [10]. Additionally, lactate uptake and oxidation in other organs may be reduced. Nevertheless, hyperlactatemia remains a predictor for mortality; however, the underlying mechanisms are possibly misunderstood. Therefore, it seems to be worth the effort to reframe the image of lactate in the mind of physiologists and clinicians. Instead of regarding lactate as a waste product, its importance as a metabolite exerting even beneficial effects in the cardiovascular system should be considered, as shown by Homilius and colleagues [4].
Cor de Wit: conceptualization, writing – original draft, writing – review and editing, visualization, funding acquisition.
期刊介绍:
Acta Physiologica is an important forum for the publication of high quality original research in physiology and related areas by authors from all over the world. Acta Physiologica is a leading journal in human/translational physiology while promoting all aspects of the science of physiology. The journal publishes full length original articles on important new observations as well as reviews and commentaries.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


