Neglecting non-vascular plants leads to underestimation of grassland plant diversity loss under experimental nutrient addition
IF 5.6
1区 环境科学与生态学
Q1 ECOLOGY
引用次数: 0
Abstract
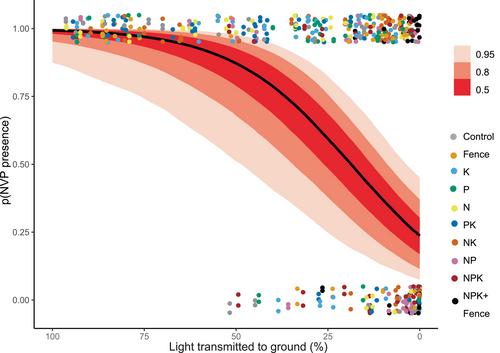




忽略非维管植物会导致对实验营养物添加下草地植物多样性损失的低估
生态学理论预测,营养物质的添加会导致植物多样性的丧失(Tilman, 1982),这已经在世界范围内的许多植物群落研究中得到了实验证明(Borer, Seabloom, et ., 2014;Harpole et al., 2016;Seabloom, Batzer等,2021)。此外,理论和实证研究表明,当食草动物增加地面光照时,营养引起的植物多样性损失可能会得到缓解(Borer, Seabloom等,2014;Olff,里奇,1998)。陆地系统植物多样性驱动因素的实验研究通常集中在维管植物(VP)上,而非维管植物(NVP;这里是苔藓植物),通常被忽略,尽管nvp通常可以代表草地群落中总植物多样性的20%-40% (Dengler等,2020;克劳斯,穆勒,2014;Löbel等,2006;Lyons et al., 2022;Tansley,亚当森,1925)。因此,目前尚不清楚NVP多样性对养分添加或放牧的响应是否与VP一致,在这种情况下VP响应可以预测NVP响应。然而,NVPs可能会有不同的反应(例如减少到或多或少的程度),在这种情况下,它们的加入将改善我们对陆地系统中植物多样性丧失的理解。在草地生态系统中,NVPs主要由陆地植物的两个分支(苔藓植物)和地衣植物)组成,它们有助于许多生态系统的功能和特性。苔藓植物通常占地上生物量的很大比例(Boch et al., 2018;Hejcman et al., 2010;Wielgolaski, 1972),调节小气候(Jaroszynska et al., 2023),水文(Michel et al., 2013),碳和养分循环(O'Neill, 2000;Turetsky, 2003),固氮作用(Lindo等,2013),并影响本地和外来维管植物的种子萌发(Dollery等,2022),土壤微生物多样性(Xiao等,2023)和土壤多功能性(Xiao等,2024)。因此,陆地系统中NVPs的存在和多样性即使在vp主导的生态系统中也会产生深远的生态影响。NVPs和vp之间的重要差异包括NVPs比vp短一个数量级,缺乏有效的传导组织、根和气孔,光合速率相对较低,这可能会影响它们的生态作用(Rydin, 2009)。它们的矮小身材使非能见度能见度能见度较差,因此,当营养供应增加加剧了对光线的竞争时,非能见度能见度可能比非能见度能见度更高(Rydin, 2009)。在过去的几十年里,一些实证研究提供了NVP对增加的养分可用性或放牧压力的反应的预期。实验营养物添加可以降低不同生态系统中NVPs的多样性(Bergamini &;泡利,2001;Jägerbrand等,2006;Jappinen,Hotanen, 1990;约,1992;威尔逊,Tilman, 2002),通常是由于与副总裁对光的竞争加剧(Aude &;Ejrnæ年代,2005;Bergamini,泡利,2001;Boch et al., 2018;Cusell et al., 2014;约,1992;van der Wal et al., 2005;Virtanen等人,2017)。过多的营养负荷也可能对NVP的生存产生直接的负面影响(Armitage et al., 2012;皮尔斯等人,2003;Virtanen et al., 2000)。虽然氮(N)通常被认为是影响NVPs最重要的养分,但氮和磷(P)的联合添加比单一养分添加对NVPs的负面影响更大(Bergamini &;泡利,2001;Øien et al., 2018)。然而,营养素的影响在研究中并不完全一致,一些研究发现对NVPs的影响相对较弱(Stevens等人,2004,2006;Sun et al., 2017),甚至营养水平与NVP丰度或物种多样性之间存在正相关关系(Gordon et al., 2001;Ingerpuu等人,1998;Slavik et al., 2004)。因此,NVPs对营养添加的反应是可变的,这表明这些反应与环境有关。与营养效应一样,放牧对NVPs的影响也有积极的影响(Bernes等人,2018;Chollet et al., 2013;Ingerpuu,收,2015;Takala et al., 2012, 2014;Tansley,Adamson, 1925),中性(Austrheim et al., 2007)或负面(Boch et al., 2018;Spitale, 2021;维尔塔宁,克劳利,2010)。草食动物可以通过促进当地定植、刺激土壤一水硬铝石库发芽或改变植物竞争相互作用来增强植物多样性(Olff &;里奇,1998)。草食动物去除生物量可以减轻VPs对NVPs的竞争效应,补偿施肥的负面影响(Aude &;Ejrnæ年代,2005)。另一方面,强烈的放牧也可能作为一种强烈的干扰,导致NVP物种的损失(Downing, 1992)。 基于现有的、稀缺的和混合的证据,NVP多样性如何单独响应养分添加和放牧压力的变化以及相互作用仍然不确定。重要的是,如果NVPs的反应不同于vp,这可能会改变对草地植物多样性损失率的预测。忽略nvp可能导致在养分添加和/或放牧变化下物种损失率的高估或低估。更具体地说,这些结果可能在下列条件下产生:(i)对物种损失率估计没有影响。如果NVP和VP对营养添加和放牧排斥的反应相似,或者NVP太稀缺,多样性太低而无法产生任何影响,则会出现这种情况。高估物种损失率。如果增加营养物质或改变放牧水平所带来的直接NVP多样性效益,可以通过增加NVP多样性来补偿vp的损失,则可能发生这种情况。低估物种损失率。如果NVP多样性的下降幅度大于VP多样性,则可能发生这种情况,原因是营养物质直接抑制了NVP,或者由于与VP争夺光线的竞争加剧。在这里,我们测试了营养添加和放牧排除对NVP和VP多样性的影响,这些站点是全球分布的营养网络实验的一部分(Borer, Harpole, et al., 2014)。这些地点在物种组成和环境背景上存在差异,这使得研究人员能够找到驱动植物多样性的反应和机制的共性。具体而言,我们比较了NVP和VP物种丰富度、多样性和均匀性对不同营养物质(N、P、K)的因子添加以及对NPK+μ和放牧排除处理的响应,以揭示考虑NVP的响应是否会改变目前仅基于vvp的草地植物多样性变化的总体估计。
本文章由计算机程序翻译,如有差异,请以英文原文为准。
求助全文
约1分钟内获得全文
求助全文
来源期刊

Journal of Ecology
环境科学-生态学
CiteScore
10.90
自引率
5.50%
发文量
207
审稿时长
3.0 months
期刊介绍:
Journal of Ecology publishes original research papers on all aspects of the ecology of plants (including algae), in both aquatic and terrestrial ecosystems. We do not publish papers concerned solely with cultivated plants and agricultural ecosystems. Studies of plant communities, populations or individual species are accepted, as well as studies of the interactions between plants and animals, fungi or bacteria, providing they focus on the ecology of the plants.
We aim to bring important work using any ecological approach (including molecular techniques) to a wide international audience and therefore only publish papers with strong and ecological messages that advance our understanding of ecological principles.
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


