Ellen O. Aikens, Jessica Speiser, Karma Choki, Michele Lovara, Anna Weesies, Jeffrey Tillery, Sean Ryder, Erica Lafferty, Amanda E. Cheeseman, William J. Severud, Hall Sawyer
{"title":"Challenging conventional views on the elevational limits of pronghorn habitat","authors":"Ellen O. Aikens, Jessica Speiser, Karma Choki, Michele Lovara, Anna Weesies, Jeffrey Tillery, Sean Ryder, Erica Lafferty, Amanda E. Cheeseman, William J. Severud, Hall Sawyer","doi":"10.1002/ecy.4422","DOIUrl":null,"url":null,"abstract":"<p>Innovations in animal tracking technologies have allowed researchers to gain an unprecedented view into the daily lives of animals, advancing our understanding of their ecology, evolution, and physiology (Kays et al., <span>2015</span>), while also providing insights that can defy conventional knowledge. For example, lightweight tracking devices revealed that migratory moths (<i>Acherontia atropos</i>) actively navigate, calling into question the widespread assumption that invertebrate migration is largely driven by passive movement dictated by prevailing winds (Menz et al., <span>2022</span>). Here, we present another example that challenges conventional knowledge of what constitutes typical pronghorn (<i>Antilocapra americana</i>) habitat by highlighting the use of extraordinary high-elevation summer ranges in the Carter Mountain area within the Greater Yellowstone Ecosystem (GYE) in Wyoming, USA (hereafter the Carter Mountain area; Figure 1). Furthermore, this discovery of high-elevation habitat use, which only occurs in a portion of the population, raises questions about the processes that generate and maintain diversity in animal movement tactics across individuals that share a common winter range and have access to the same resources (Lundberg, <span>1988</span>; Table 1).</p><p>Pronghorn are the only extant member of the family Antilocapridae, a species that is known for remarkable speed, endurance, and eyesight (O'Gara & Yoakum, <span>2004</span>). Although the geographic range of pronghorn spans from Canada to Mexico, the extent of pronghorn habitat is generally restricted to open basins and shrublands with flat or gentle topography (O'Gara & Yoakum, <span>2004</span>; Yoakum, <span>1981</span>). Such habitat preferences enhance the ability of pronghorn to detect, avoid, and evade predators (Berger et al., <span>1983</span>; O'Gara & Yoakum, <span>2004</span>). Notably, these habitats typically occur in arid or semiarid regions ranging from ~900 to 1800 m in elevation (O'Gara & Yoakum, <span>2004</span>; Yoakum, <span>1981</span>). Pronghorn are also sensitive to human disturbances, avoiding roads, suburban development, and industrial development (Gavin & Komers, <span>2006</span>; Reinking et al., <span>2019</span>; Sandoval Lambert et al., <span>2022</span>). Beyond static landscape features—such as elevation or habitat type—another key habitat requirement for pronghorn is the ability to freely move across the landscape to seek out high-quality forage during the growing season and respond to unpredictable environmental conditions (Gates et al., <span>2012</span>). To meet these seasonally-changing resource requirements, pronghorn are known for their highly flexible and wide-ranging movements, including long-distance migrations (Berger, <span>2004</span>; Reinking et al., <span>2019</span>). Thus, maintaining landscape connectivity by minimizing barriers to movement is a critical component of pronghorn conservation efforts (Gates et al., <span>2012</span>).</p><p>To delineate key movement corridors for pronghorn in the Carter Mountain area, a 2-year (November 2019 to November 2021) GPS-tracking study was initiated (<i>n</i> = 128 adult [>1.5 years], female pronghorn). GPS tracking revealed that nearly 20% of the pronghorn population gained over 1500 m in elevation, moving from lower elevation sagebrush basins (~1100–1700 m) to summer in high-elevation alpine meadows and plateaus (~2750–3400 m; Figure 1). Such a high-elevation alpine habitat above 3000 m is often assumed to be either inaccessible or unsuitable to pronghorn. Although the use of high-elevation habitat by pronghorn has previously been described anecdotally, such accounts are often considered anomalous (e.g., Thilenius, <span>1985</span>). Here, the deployment of GPS-tracking technology on pronghorn in the Carter Mountain area provides a rare and detailed glimpse into this poorly described behavior, highlighting where, when, and for how long pronghorn use high-elevation, alpine habitats.</p><p>Of the 113 GPS-collared pronghorn monitored long enough to determine migratory status, approximately half were migratory (<i>n</i> = 58), and the other half were nonmigratory (<i>n</i> = 55). Among the migratory individuals, GPS data revealed an unexpected seasonal migration in which 21 of the pronghorn moved across vast elevational gradients (>1500 m in elevation change) to spend the summer months in high-elevation alpine habitat. On average, these high-elevation migrants gained 1843 m in elevation, with some individuals gaining a remarkable 2197 m of elevation (Figure 1). When migrating to the highest-elevation regions, pronghorn often moved rapidly (see Appendix S1 for details). To quantify the diversity in movement tactics used by pronghorn in the Carter Mountain area, we classified the annual change in elevation for each animal year of tracking data as low (<500 m; 30.8% of animal years), medium (500–1500 m; 52.4% of animal years), or high (>1500 m; 16.8% of animal years; Figure 2A,B). The use of habitats above 3000 m started in late April (Day of Year [DOY] = 117), peaked in early August (DOY = 220), and ended by mid-October (DOY = 284; Figure 2C). Individuals that gained 1500 m in elevation annually spent an average of 39.3 days above 3000 m per year (range = 0–106 days; Figure 2D). Of the 17 individuals with two years of tracking data that were classified as high-elevation migrants, 65% maintained this behavior across years. The remaining 35% of high-elevation individuals (<i>n</i> = 6) that switched tactics across years spent less time above 3000 m in the previous year (mean = 14.5 days) than those individuals that remained consistent in their classification across tracking years (mean = 39.3 days). In accordance with the State Dependent Hypothesis, this switching behavior could also be explained by previous experience or the state of the individual (e.g., body condition, age, or lactation status; Table 1; Lundberg, <span>1988</span>).</p><p>To evaluate the novelty of this alpine habitat use by pronghorn in the Carter Mountain area and to assess the possible existence of similar behaviors by pronghorn in other systems, we conducted a systematic review of both the gray and primary literature. We employed Publish or Perish 8 software (Harzing, <span>2016</span>) to search titles, keywords, and text from Google Scholar, returning the 500 most relevant papers using the search terms: “migration” OR “movement” OR “summer range” OR “winter range” OR “VHF” OR “telemetry” OR “radio” AND “pronghorn” AND “elevation.” We reviewed each paper and recorded descriptions of the range of elevations used by pronghorn. We excluded reports of study area elevations that did not denote areas used by pronghorn. Papers where pronghorn were not a focal species (<i>n</i> = 150), or were otherwise nonstandardized, duplicative, or inaccessible were also excluded (<i>n</i> = 24). This resulted in 326 relevant sources, but only 57 (17.5%) of those sources reported elevation. Only four sources reported pronghorn using elevations greater than 3000 m, including mentions of populations in northern New Mexico and Wyoming (Cramer et al., <span>2022</span>; Kauffman et al,, <span>2022</span>; Yoakum, <span>1981</span>). Only recently has high-elevation habitat use been described using GPS tracking in agency reports (Kauffman et al., <span>2022</span>; Sawyer & Telander, <span>2022</span>). Based on this literature review, the analysis presented here represents the first detailed description of habitat use above 3000 m by pronghorn within the primary literature.</p><p>The use of alpine habitats raises several questions about the processes that generate and maintain this behavior, alongside the resident and low-elevation migrants in the Carter Mountain area (Table 1). Gaining up to 2197 m annually, and summering at such high elevations requires a long (64–97 km; Sawyer & Telander, <span>2022</span>) and potentially difficult migration that may not be possible for all individuals (sensu the State Dependent and Bet Hedging Hypotheses; Table 1). Summering at high elevations presumably exposes pronghorn to vastly different predation pressures and forage composition compared with the lower elevation basins that typically characterize pronghorn habitat. While the benefits of delayed vegetation phenology associated with elevation and moisture gradients are well documented for migratory ungulates (Aikens et al., <span>2017</span>), the diet composition and preferences of pronghorn in alpine habitats is generally unknown. The higher elevation regions of the GYE also support a full suite of large carnivores, including grizzly bears (<i>Ursus arctos</i>) and wolves (<i>Canis lupus</i>). Likewise, high-elevation habitats also host a wider diversity of grazing species in the summer (e.g., cattle [<i>Bos</i> spp.], elk [<i>Cervus elaphus</i>], mule deer [<i>Odocoileus hemionus</i>] and bighorn sheep [<i>Ovis canadensis</i>]) in comparison with lower elevation regions that likely have less interspecific competition but more intraspecific competition. It remains unclear how pronghorn balance the risks and rewards of living in alpine environments that support such a diverse predator assemblage (sensu the Fitness Balancing Hypothesis) as well as heterogeneity in competition within and across species (sensu the Frequency-Dependent Selection Hypothesis; Table 1). For example, do high-elevation pronghorn accept the risk of increased predation pressure to capitalize on abundant forage? Or can alpine pronghorn modify their behavior to mitigate risks by increasing the size of foraging groups or shifting activity levels to minimize predator encounter rates? Future research comparing the group sizes and activity budgets of pronghorn summering at high, mid, and low elevations could provide insights into how differences in forage availability and predation shapes the social structure and activity patterns of pronghorn. Collecting long-term data on group sizes, activity, individual state, and fitness metrics (i.e., survival and reproduction) across the full range of movement tactics in this system could also provide insight into the processes that generate and maintain this diversity, allowing for tests of the hypotheses listed in Table 1.</p><p>Beyond understanding how pronghorn use alpine environments, the existence of these high-elevation migrants also raises questions about how this behavior developed, how long it has persisted on the landscape, and the role that such a specialized movement tactic plays in the broader population dynamics of this pronghorn herd. Migratory behaviors can take decades to establish (Jesmer et al., <span>2018</span>) and presumably even longer for migrations that involve moving long distances across rugged terrain. We found one account from a 1985 Forest Service report noting the “surprising” presence of pronghorn in the alpine zone of the Carter Mountains (Thilenius, <span>1985</span>), suggesting that this high-elevation subset of the population has likely persisted for at least 40 years, prior to large carnivore restoration. Although we did not examine the archaeological records, we suspect that such investigations may be a useful source for reconstructing pronghorn use of alpine habitats. Regardless of how or when this migration was established, the conservation of migratory routes and the maintenance of diverse movement tactics within a population likely increases the abundance, resilience, and long-term viability of ungulate populations (Lowrey et al., <span>2020</span>). While much remains unclear about the prevalence, development, and life history of pronghorn utilizing alpine habitat, our work suggests that the importance of high-elevation ranges to pronghorn are not limited to the GYE (Kauffman et al., <span>2022</span>), and that our conventional views on the elevational limits—and associated habitats—of pronghorn should be broadened.</p><p>The authors declare no conflicts of interest.</p>","PeriodicalId":11484,"journal":{"name":"Ecology","volume":"105 11","pages":""},"PeriodicalIF":4.4000,"publicationDate":"2024-09-29","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1002/ecy.4422","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"Ecology","FirstCategoryId":"93","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1002/ecy.4422","RegionNum":2,"RegionCategory":"环境科学与生态学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"ECOLOGY","Score":null,"Total":0}
引用次数: 0
Abstract
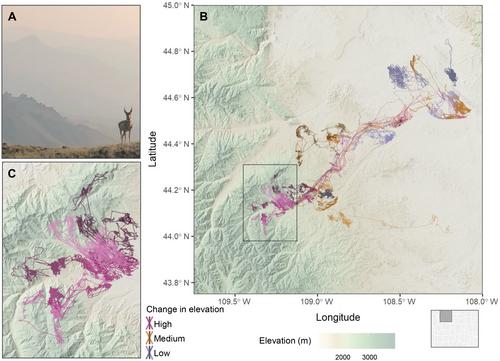
Innovations in animal tracking technologies have allowed researchers to gain an unprecedented view into the daily lives of animals, advancing our understanding of their ecology, evolution, and physiology (Kays et al., 2015), while also providing insights that can defy conventional knowledge. For example, lightweight tracking devices revealed that migratory moths (Acherontia atropos) actively navigate, calling into question the widespread assumption that invertebrate migration is largely driven by passive movement dictated by prevailing winds (Menz et al., 2022). Here, we present another example that challenges conventional knowledge of what constitutes typical pronghorn (Antilocapra americana) habitat by highlighting the use of extraordinary high-elevation summer ranges in the Carter Mountain area within the Greater Yellowstone Ecosystem (GYE) in Wyoming, USA (hereafter the Carter Mountain area; Figure 1). Furthermore, this discovery of high-elevation habitat use, which only occurs in a portion of the population, raises questions about the processes that generate and maintain diversity in animal movement tactics across individuals that share a common winter range and have access to the same resources (Lundberg, 1988; Table 1).
Pronghorn are the only extant member of the family Antilocapridae, a species that is known for remarkable speed, endurance, and eyesight (O'Gara & Yoakum, 2004). Although the geographic range of pronghorn spans from Canada to Mexico, the extent of pronghorn habitat is generally restricted to open basins and shrublands with flat or gentle topography (O'Gara & Yoakum, 2004; Yoakum, 1981). Such habitat preferences enhance the ability of pronghorn to detect, avoid, and evade predators (Berger et al., 1983; O'Gara & Yoakum, 2004). Notably, these habitats typically occur in arid or semiarid regions ranging from ~900 to 1800 m in elevation (O'Gara & Yoakum, 2004; Yoakum, 1981). Pronghorn are also sensitive to human disturbances, avoiding roads, suburban development, and industrial development (Gavin & Komers, 2006; Reinking et al., 2019; Sandoval Lambert et al., 2022). Beyond static landscape features—such as elevation or habitat type—another key habitat requirement for pronghorn is the ability to freely move across the landscape to seek out high-quality forage during the growing season and respond to unpredictable environmental conditions (Gates et al., 2012). To meet these seasonally-changing resource requirements, pronghorn are known for their highly flexible and wide-ranging movements, including long-distance migrations (Berger, 2004; Reinking et al., 2019). Thus, maintaining landscape connectivity by minimizing barriers to movement is a critical component of pronghorn conservation efforts (Gates et al., 2012).
To delineate key movement corridors for pronghorn in the Carter Mountain area, a 2-year (November 2019 to November 2021) GPS-tracking study was initiated (n = 128 adult [>1.5 years], female pronghorn). GPS tracking revealed that nearly 20% of the pronghorn population gained over 1500 m in elevation, moving from lower elevation sagebrush basins (~1100–1700 m) to summer in high-elevation alpine meadows and plateaus (~2750–3400 m; Figure 1). Such a high-elevation alpine habitat above 3000 m is often assumed to be either inaccessible or unsuitable to pronghorn. Although the use of high-elevation habitat by pronghorn has previously been described anecdotally, such accounts are often considered anomalous (e.g., Thilenius, 1985). Here, the deployment of GPS-tracking technology on pronghorn in the Carter Mountain area provides a rare and detailed glimpse into this poorly described behavior, highlighting where, when, and for how long pronghorn use high-elevation, alpine habitats.
Of the 113 GPS-collared pronghorn monitored long enough to determine migratory status, approximately half were migratory (n = 58), and the other half were nonmigratory (n = 55). Among the migratory individuals, GPS data revealed an unexpected seasonal migration in which 21 of the pronghorn moved across vast elevational gradients (>1500 m in elevation change) to spend the summer months in high-elevation alpine habitat. On average, these high-elevation migrants gained 1843 m in elevation, with some individuals gaining a remarkable 2197 m of elevation (Figure 1). When migrating to the highest-elevation regions, pronghorn often moved rapidly (see Appendix S1 for details). To quantify the diversity in movement tactics used by pronghorn in the Carter Mountain area, we classified the annual change in elevation for each animal year of tracking data as low (<500 m; 30.8% of animal years), medium (500–1500 m; 52.4% of animal years), or high (>1500 m; 16.8% of animal years; Figure 2A,B). The use of habitats above 3000 m started in late April (Day of Year [DOY] = 117), peaked in early August (DOY = 220), and ended by mid-October (DOY = 284; Figure 2C). Individuals that gained 1500 m in elevation annually spent an average of 39.3 days above 3000 m per year (range = 0–106 days; Figure 2D). Of the 17 individuals with two years of tracking data that were classified as high-elevation migrants, 65% maintained this behavior across years. The remaining 35% of high-elevation individuals (n = 6) that switched tactics across years spent less time above 3000 m in the previous year (mean = 14.5 days) than those individuals that remained consistent in their classification across tracking years (mean = 39.3 days). In accordance with the State Dependent Hypothesis, this switching behavior could also be explained by previous experience or the state of the individual (e.g., body condition, age, or lactation status; Table 1; Lundberg, 1988).
To evaluate the novelty of this alpine habitat use by pronghorn in the Carter Mountain area and to assess the possible existence of similar behaviors by pronghorn in other systems, we conducted a systematic review of both the gray and primary literature. We employed Publish or Perish 8 software (Harzing, 2016) to search titles, keywords, and text from Google Scholar, returning the 500 most relevant papers using the search terms: “migration” OR “movement” OR “summer range” OR “winter range” OR “VHF” OR “telemetry” OR “radio” AND “pronghorn” AND “elevation.” We reviewed each paper and recorded descriptions of the range of elevations used by pronghorn. We excluded reports of study area elevations that did not denote areas used by pronghorn. Papers where pronghorn were not a focal species (n = 150), or were otherwise nonstandardized, duplicative, or inaccessible were also excluded (n = 24). This resulted in 326 relevant sources, but only 57 (17.5%) of those sources reported elevation. Only four sources reported pronghorn using elevations greater than 3000 m, including mentions of populations in northern New Mexico and Wyoming (Cramer et al., 2022; Kauffman et al,, 2022; Yoakum, 1981). Only recently has high-elevation habitat use been described using GPS tracking in agency reports (Kauffman et al., 2022; Sawyer & Telander, 2022). Based on this literature review, the analysis presented here represents the first detailed description of habitat use above 3000 m by pronghorn within the primary literature.
The use of alpine habitats raises several questions about the processes that generate and maintain this behavior, alongside the resident and low-elevation migrants in the Carter Mountain area (Table 1). Gaining up to 2197 m annually, and summering at such high elevations requires a long (64–97 km; Sawyer & Telander, 2022) and potentially difficult migration that may not be possible for all individuals (sensu the State Dependent and Bet Hedging Hypotheses; Table 1). Summering at high elevations presumably exposes pronghorn to vastly different predation pressures and forage composition compared with the lower elevation basins that typically characterize pronghorn habitat. While the benefits of delayed vegetation phenology associated with elevation and moisture gradients are well documented for migratory ungulates (Aikens et al., 2017), the diet composition and preferences of pronghorn in alpine habitats is generally unknown. The higher elevation regions of the GYE also support a full suite of large carnivores, including grizzly bears (Ursus arctos) and wolves (Canis lupus). Likewise, high-elevation habitats also host a wider diversity of grazing species in the summer (e.g., cattle [Bos spp.], elk [Cervus elaphus], mule deer [Odocoileus hemionus] and bighorn sheep [Ovis canadensis]) in comparison with lower elevation regions that likely have less interspecific competition but more intraspecific competition. It remains unclear how pronghorn balance the risks and rewards of living in alpine environments that support such a diverse predator assemblage (sensu the Fitness Balancing Hypothesis) as well as heterogeneity in competition within and across species (sensu the Frequency-Dependent Selection Hypothesis; Table 1). For example, do high-elevation pronghorn accept the risk of increased predation pressure to capitalize on abundant forage? Or can alpine pronghorn modify their behavior to mitigate risks by increasing the size of foraging groups or shifting activity levels to minimize predator encounter rates? Future research comparing the group sizes and activity budgets of pronghorn summering at high, mid, and low elevations could provide insights into how differences in forage availability and predation shapes the social structure and activity patterns of pronghorn. Collecting long-term data on group sizes, activity, individual state, and fitness metrics (i.e., survival and reproduction) across the full range of movement tactics in this system could also provide insight into the processes that generate and maintain this diversity, allowing for tests of the hypotheses listed in Table 1.
Beyond understanding how pronghorn use alpine environments, the existence of these high-elevation migrants also raises questions about how this behavior developed, how long it has persisted on the landscape, and the role that such a specialized movement tactic plays in the broader population dynamics of this pronghorn herd. Migratory behaviors can take decades to establish (Jesmer et al., 2018) and presumably even longer for migrations that involve moving long distances across rugged terrain. We found one account from a 1985 Forest Service report noting the “surprising” presence of pronghorn in the alpine zone of the Carter Mountains (Thilenius, 1985), suggesting that this high-elevation subset of the population has likely persisted for at least 40 years, prior to large carnivore restoration. Although we did not examine the archaeological records, we suspect that such investigations may be a useful source for reconstructing pronghorn use of alpine habitats. Regardless of how or when this migration was established, the conservation of migratory routes and the maintenance of diverse movement tactics within a population likely increases the abundance, resilience, and long-term viability of ungulate populations (Lowrey et al., 2020). While much remains unclear about the prevalence, development, and life history of pronghorn utilizing alpine habitat, our work suggests that the importance of high-elevation ranges to pronghorn are not limited to the GYE (Kauffman et al., 2022), and that our conventional views on the elevational limits—and associated habitats—of pronghorn should be broadened.
期刊介绍:
Ecology publishes articles that report on the basic elements of ecological research. Emphasis is placed on concise, clear articles documenting important ecological phenomena. The journal publishes a broad array of research that includes a rapidly expanding envelope of subject matter, techniques, approaches, and concepts: paleoecology through present-day phenomena; evolutionary, population, physiological, community, and ecosystem ecology, as well as biogeochemistry; inclusive of descriptive, comparative, experimental, mathematical, statistical, and interdisciplinary approaches.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


