{"title":"Phytotelmata-dwelling frog larvae might exhibit no defecation: A unique adaptation to a closed aquatic environment","authors":"Bun Ito, Yasukazu Okada","doi":"10.1002/ecy.4428","DOIUrl":null,"url":null,"abstract":"<p>The Taiwanese tree frog <i>Kurixalus eiffingeri</i> lays its eggs in small water pools on plants, such as tree hollows or bamboo stumps (i.e., phytotelmata, Figure 1a,b). The larvae feed exclusively on the trophic eggs laid by their mothers and spend their entire larval period in tiny phytotelmata of less than 50 cm<sup>2</sup> of surface area and 2.5 cm of water depth (Chen et al., <span>2007</span>; Kam et al., <span>1997</span>, <span>2000</span>; Sato & Iwai, <span>2021</span>; Ueda, <span>1986</span>). Interestingly, during laboratory rearing, we observed that <i>K. eiffingeri</i> larvae do not seem to defecate until metamorphosis (Figure 1c), and that larvae accumulate solid feces in their gut during whole larval period (Figure 1d,e). From this finding, we hypothesized that this is a unique sanitation strategy to avoid potentially harmful ammonia contamination in a small, closed, water system.</p><p>Animals excrete nitrogen compounds when they metabolize protein nutrient sources. The mode of nitrogen excretion of animals varies according to their living environments. Generally, aquatic teleost fish and amphibian larvae metabolize waste nitrogen compounds to ammonia which is then excreted (Wright, <span>1995</span>). The large amount of water around aquatic animals quickly dilutes the excreted ammonia and prevents ammonia poisoning (Wright et al., <span>1993</span>). On the other hand, adult terrestrial amphibians and mammals metabolize waste nitrogen compounds to non-toxic urea which is excreted. This is because the amount of available water on land is too scarce to detoxify ammonia through dilution (Atkinson, <span>1992</span>). Similarly, reptiles and birds, whose embryos develop in hard eggshells, excrete nitrogen as uric acid, which is insoluble in water and less harmful (Wright, <span>1995</span>).</p><p>Thus, there is a close relationship between the organismal habitat and its mode of nitrogen excretion. Notably, animals living in a closed aquatic habitat may face the deleterious toxicity of nitrogen excretion. Amphibian larvae usually excrete nitrogen compounds in the form of ammonia (Wright, <span>1995</span>). However, in closed aquatic systems, ammonia may accumulate in living water, and larvae living in these environments could be exposed to high, toxic, ammonia levels. Here we show that <i>K. eiffingeri</i> larvae show unique adaptation to ammonia toxicity by not excreting feces and by increasing the tolerance to ammonia.</p><p>In this study, we firstly demonstrated reduced ammonia excretion by comparing the ammonia production of <i>K. eiffingeri</i> larvae with other pond—living frog larvae (<i>Dryophytes japonicus</i>, <i>Rana ornativentris</i>, and <i>Zhangixalus arboreus</i>). Secondly, we experimentally exposed the larvae to ammonium chloride solution and showed that <i>K. eiffingeri</i> larvae have greater ammonia tolerance.</p><p><i>K. eiffingeri</i> (Anura Rhacophoridae) is a small frog (3–4 cm in adult length) which inhabits Ishigaki Island, Iriomote Island, and Taiwan. This species shows unique caretaking behavior. Amplexus and oviposition occur in a narrow water pool (less than 50 cm<sup>2</sup> of surface area and 2.5 cm of water depth) in a tree hollow or a bamboo stump (Sato & Iwai, <span>2021</span>), and the hatched larvae grow up in these phytotelmata. The time from hatching to metamorphosis varies greatly depending on the nutritional status of the larvae and takes approximately 21–43 days. The total body length of the larvae at hatching is 9.73 ± 2.35 mm (<i>N</i> = 17) and is 29.74 ± 1.13 mm (<i>N</i> = 18) when metamorphosis begins and they land (Kishimoto & Hayashi, <span>2017</span>; Ito, personal observation). After hatching, the female takes care of her offspring by feeding them with the trophic eggs laid by herself. Until metamorphosis, the larvae exclusively feed on these trophic eggs (Chen et al., <span>2007</span>; Kam et al., <span>1997</span>, <span>2000</span>; Ueda, <span>1986</span>).</p><p>Sixteen <i>K. eiffingeri</i> adults were collected from various locations in Ishigaki City, Okinawa, Japan, in May, November 2022, and May 2023. Several pairs of a female and a male were kept in terrariums with soil and plants with a volume of approximately 7 L (153 × 278 × 165 mm) and watered with a rainfall device for 1 h/day (Appendix S1: Figures S1 and S2). Light:Dark conditions were 12 h:12 h, and the temperature was kept in 24 ± 1°C. Adults were fed Turkestan cockroaches <i>Blatta (Shelfordella) lateralis</i> and frozen house crickets <i>Acheta domesticus</i> three times a week. Calcium and vitamin powders were sprinkled on the food once a week to avoid metabolic disorders, including rachitis.</p><p>Plastic cups with a base area of approximately 13 cm<sup>2</sup> were placed in the terrarium (Appendix S1: Figure S2), and the larvae were hatched and grew in water puddles inside them. The larvae fed on the maternally provided trophic eggs.</p><p>As controls for <i>K. eiffingeri</i>, larvae of Japanese tree frog <i>D. japonicus</i>, Japanese mountain brown frog <i>R. ornativentris</i>, and Japanese forest green tree frog <i>Z. arboreus</i> were also collected from the wild in Hachioji, Tokyo, Japan, from June to August 2023. Larvae of these species inhabit open-water ponds. Larvae of these three species were collected a week or less before being used in the experiment and reared under light/dark and temperature conditions in the same way as <i>K. eiffingeri</i>, as described above. Larvae of these three species were kept in 5-L containers with water, and rearing water was changed every two days. All larvae were between Gosner stages 30 to 40. Before all experiments, <i>K. eiffingeri</i> larvae were fed trophic eggs, and the other three species were fed fish food ad libitum (Seria Co., Ltd., Gifu, Japan).</p><p>The ammonia concentration in the rearing water was measured using the larvae of <i>K. eiffingeri</i> (<i>n</i> = 15), <i>D. japonicus</i> (<i>n</i> = 15), <i>R. ornativentris</i> (<i>n</i> = 15), and <i>Z. arboreus</i> (<i>n</i> = 5). We weighed all larvae before the experiment, and experimental larvae were reared in 20 mL of distilled water (DW) in a plastic case with a diameter of 3.5 cm (Figure 1f). The experiment was conducted from June to July 2023. To avoid pollution from food, larvae were not fed during this experiment. The plastic case was covered with a lid to prevent changes in ammonia concentration due to water evaporation.</p><p>We reared larvae for 9 days and ammonia concentrations in the rearing water were measured every 3 days (days 0, 3, 6, and 9). We measured the level of dissolved ammonia by Urea/Ammonia Assay Kit (Rapid) (Megazyme, Bray, Ireland) following its protocol, with NanoDrop OneC (Thermo Fisher Scientific, Massachusetts, USA) for wavelength quantification. This assay kit can measure concentrations with 0.018 mg/L as the minimum detectable level. Five microliter samples of rearing water were collected and used for measurements. Since this is a very small amount (0.00025% of total water), the collection of breeding water may not affect the ammonia concentration. In order to account for the body size difference, ammonia concentrations were corrected by individual body weights (concentration [in grams per liter]/body weight [in grams]).</p><p>Next, we measured the amount of ammonia in gut. <i>K. eiffingeri</i> was fed with trophic eggs and the other two species were fed with fish food ad libitum, then the feces-containing gut (rectum and large intestine) was taken out from the larva by dissection (Figure 1e). After removing the surface water with paper, we weighed the gut and placed it in microtubes. The gut and its contents were then homogenized and suspended in 100 μL of DW. The ammonia concentration of the suspension was measured using the same protocol as described above. This experiment was performed in August 2023 using <i>K. eiffingeri</i> (<i>n</i> = 10), <i>D. japonicus</i> (<i>n</i> = 10), and <i>R. ornativentris</i> (<i>n</i> = 5).</p><p>Larval tolerance to ammonia was experimentally tested using <i>K. eiffingeri</i> (<i>n</i> = 30) and <i>D. japonicus</i> (<i>n</i> = 30) in August 2023. Ammonium chloride is efficient to test the effect of dissolved ammonia on larval survival (Schuytema & Nebeker, <span>1999</span>). Thirty larvae of each species were individually allocated to three treatments: DW, 50, and 100 mM of ammonium chloride solutions (<i>n</i> = 10 for each, ammonium chloride, FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan). The experimental larvae were checked every 12 h for 7 days for their survival.</p><p>Survival rates of each group were compared by Kaplan–Meier curves and log-rank test, followed by Bonferroni correction for multiple comparisons.</p><p>The ammonia concentration of the rearing water was significantly higher except for the comparison with day-6 <i>R. ornativentris</i> (Figure 2a). On the whole, there was a clear pattern that <i>K. eiffingeri</i> produces a very low amount of ammonia in rearing water compared with other species.</p><p>The ammonia concentration of the larval gut was significantly higher in <i>K. eiffingeri</i> than in the other two species (Figure 2b, Dunnet's test; <i>p</i> values provided in figure caption) suggesting that <i>K. eiffingeri</i> retain ammonia in their gut during the larval period.</p><p>In distilled water, larvae of <i>D. japonicus</i> could not survive more than 7 days (Figure 2c, light gray). In contrast, all larvae of <i>K. eiffingeri</i> survived in distilled water, suggesting that <i>K. eiffingeri</i> is tolerant to the ammonia produced by themselves (Figure 2c, pale light blue). At 50 mM NH<sub>4</sub>Cl treatment, the majority of <i>K. eiffingeri</i> larvae (8/10, Figure 2c, light blue) survived for 7 days but all <i>D. japonicus</i> larvae died in 7 days (Figure 2c, gray). At the greatest dose of ammonia (100 mM NH<sub>4</sub>Cl), both <i>K. eiffingeri</i> and <i>D. japonicus</i> larvae could not survive for 7 days (Figure 2c, blue and black), implying that a high dose of ammonia is also toxic to <i>K. eiffingeri</i>.</p><p><i>K. eiffingeri</i> larvae, which grow in closed system tree and stump holes, excrete less ammonia into their environment than other species whose habitats are open water ponds; they also retain high concentrations of ammonia in their gut (Figure 2a,b). This is an indirect indication that <i>K. eiffingeri</i> larvae grow without, or at least reduced, defecation. Additionally, <i>K. eiffingeri</i> larvae were shown to be more tolerant to ammonia than <i>D. japonicus</i> (Figure 2c). These results indicate that the larvae of this species have evolved a unique sanitation strategy to hold the ammonia-containing feces in the larval gut and an unusual tolerance against environmental ammonia. However, <i>K. eiffingeri</i> larvae do emit ammonia, albeit in small amounts, and although the authors have observed that <i>K. eiffingeri</i> larvae do not defecate solid feces, it is possible that they emit ammonia in liquid or other forms, which is an issue that needs further investigation.</p><p>For growth and somatic maintenance, animals must take proteinous diets, and consequently, nitrogen excretion is inevitable. Some animals may suffer from the toxic pollution of nitrogen-rich feces. <i>K. eiffingeri</i> firstly defecates just after metamorphosis to adults (Ito, personal observation). This suggests that the nitrogen brought into the phytothermata is eventually carried outside. Females of this species sequentially use the same phytothermata as a breeding site, and as a result, siblings with the same and different clutches grow together and/or sequentially in the same phytothermata (Kam et al., <span>2000</span>; Ito, personal observation). In this respect, individual sanitation behavior can also work as a social sanitation that eventually raises the fitness of siblings by keeping the growth habitat clean. Such social aspects of sanitation behavior parallel those observed in social insects where larvae of some bees and ants retain their feces in the gut until metamorphosis, bringing the social sanitation benefit to their nestmates (Jackson & Hart, <span>2009</span>). Growth in narrow, crowding environments with conspecifics, especially siblings, may be a common ecological factor that drives the evolution of the social sanitation behavior.</p><p>The question also arises whether similar phenomena can be observed in organisms living in phytotelmata. For example, many species of poison dart frogs (Dendrobatidae) also grow their larvae in phytotelmata (Lima et al., <span>2007</span>; Summers, <span>1990</span>; Weygoldt, <span>1987</span>). We predict that these species may also have similar sanitation strategies. Besides anurans, species that utilize phytotelmata are known in a very wide range of taxa, from protozoa, invertebrates, and vertebrates (Frank, <span>1983</span>). It will be an important future question to see whether the sanitation adaptation and the tolerances against ammonia are widely observed in various taxonomic groups in this extremely tiny microenvironment. This study has shed light on the novel sanitation behavior and unique biological material flow in phytotelmata, the less conspicuous but frequent and important inland aquatic system for animals (Frank, <span>1983</span>).</p><p>The authors declare no conflicts of interest.</p><p>These species are not regulated by the Tokyo Metropolitan University ethics committee or protected by Japanese law. Thus, no ethics approval was required to declare, and no special permits were needed to conduct this research.</p>","PeriodicalId":11484,"journal":{"name":"Ecology","volume":"105 11","pages":""},"PeriodicalIF":4.3000,"publicationDate":"2024-09-22","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1002/ecy.4428","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"Ecology","FirstCategoryId":"93","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1002/ecy.4428","RegionNum":2,"RegionCategory":"环境科学与生态学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"ECOLOGY","Score":null,"Total":0}
引用次数: 0
Abstract
The Taiwanese tree frog Kurixalus eiffingeri lays its eggs in small water pools on plants, such as tree hollows or bamboo stumps (i.e., phytotelmata, Figure 1a,b). The larvae feed exclusively on the trophic eggs laid by their mothers and spend their entire larval period in tiny phytotelmata of less than 50 cm2 of surface area and 2.5 cm of water depth (Chen et al., 2007; Kam et al., 1997, 2000; Sato & Iwai, 2021; Ueda, 1986). Interestingly, during laboratory rearing, we observed that K. eiffingeri larvae do not seem to defecate until metamorphosis (Figure 1c), and that larvae accumulate solid feces in their gut during whole larval period (Figure 1d,e). From this finding, we hypothesized that this is a unique sanitation strategy to avoid potentially harmful ammonia contamination in a small, closed, water system.
Animals excrete nitrogen compounds when they metabolize protein nutrient sources. The mode of nitrogen excretion of animals varies according to their living environments. Generally, aquatic teleost fish and amphibian larvae metabolize waste nitrogen compounds to ammonia which is then excreted (Wright, 1995). The large amount of water around aquatic animals quickly dilutes the excreted ammonia and prevents ammonia poisoning (Wright et al., 1993). On the other hand, adult terrestrial amphibians and mammals metabolize waste nitrogen compounds to non-toxic urea which is excreted. This is because the amount of available water on land is too scarce to detoxify ammonia through dilution (Atkinson, 1992). Similarly, reptiles and birds, whose embryos develop in hard eggshells, excrete nitrogen as uric acid, which is insoluble in water and less harmful (Wright, 1995).
Thus, there is a close relationship between the organismal habitat and its mode of nitrogen excretion. Notably, animals living in a closed aquatic habitat may face the deleterious toxicity of nitrogen excretion. Amphibian larvae usually excrete nitrogen compounds in the form of ammonia (Wright, 1995). However, in closed aquatic systems, ammonia may accumulate in living water, and larvae living in these environments could be exposed to high, toxic, ammonia levels. Here we show that K. eiffingeri larvae show unique adaptation to ammonia toxicity by not excreting feces and by increasing the tolerance to ammonia.
In this study, we firstly demonstrated reduced ammonia excretion by comparing the ammonia production of K. eiffingeri larvae with other pond—living frog larvae (Dryophytes japonicus, Rana ornativentris, and Zhangixalus arboreus). Secondly, we experimentally exposed the larvae to ammonium chloride solution and showed that K. eiffingeri larvae have greater ammonia tolerance.
K. eiffingeri (Anura Rhacophoridae) is a small frog (3–4 cm in adult length) which inhabits Ishigaki Island, Iriomote Island, and Taiwan. This species shows unique caretaking behavior. Amplexus and oviposition occur in a narrow water pool (less than 50 cm2 of surface area and 2.5 cm of water depth) in a tree hollow or a bamboo stump (Sato & Iwai, 2021), and the hatched larvae grow up in these phytotelmata. The time from hatching to metamorphosis varies greatly depending on the nutritional status of the larvae and takes approximately 21–43 days. The total body length of the larvae at hatching is 9.73 ± 2.35 mm (N = 17) and is 29.74 ± 1.13 mm (N = 18) when metamorphosis begins and they land (Kishimoto & Hayashi, 2017; Ito, personal observation). After hatching, the female takes care of her offspring by feeding them with the trophic eggs laid by herself. Until metamorphosis, the larvae exclusively feed on these trophic eggs (Chen et al., 2007; Kam et al., 1997, 2000; Ueda, 1986).
Sixteen K. eiffingeri adults were collected from various locations in Ishigaki City, Okinawa, Japan, in May, November 2022, and May 2023. Several pairs of a female and a male were kept in terrariums with soil and plants with a volume of approximately 7 L (153 × 278 × 165 mm) and watered with a rainfall device for 1 h/day (Appendix S1: Figures S1 and S2). Light:Dark conditions were 12 h:12 h, and the temperature was kept in 24 ± 1°C. Adults were fed Turkestan cockroaches Blatta (Shelfordella) lateralis and frozen house crickets Acheta domesticus three times a week. Calcium and vitamin powders were sprinkled on the food once a week to avoid metabolic disorders, including rachitis.
Plastic cups with a base area of approximately 13 cm2 were placed in the terrarium (Appendix S1: Figure S2), and the larvae were hatched and grew in water puddles inside them. The larvae fed on the maternally provided trophic eggs.
As controls for K. eiffingeri, larvae of Japanese tree frog D. japonicus, Japanese mountain brown frog R. ornativentris, and Japanese forest green tree frog Z. arboreus were also collected from the wild in Hachioji, Tokyo, Japan, from June to August 2023. Larvae of these species inhabit open-water ponds. Larvae of these three species were collected a week or less before being used in the experiment and reared under light/dark and temperature conditions in the same way as K. eiffingeri, as described above. Larvae of these three species were kept in 5-L containers with water, and rearing water was changed every two days. All larvae were between Gosner stages 30 to 40. Before all experiments, K. eiffingeri larvae were fed trophic eggs, and the other three species were fed fish food ad libitum (Seria Co., Ltd., Gifu, Japan).
The ammonia concentration in the rearing water was measured using the larvae of K. eiffingeri (n = 15), D. japonicus (n = 15), R. ornativentris (n = 15), and Z. arboreus (n = 5). We weighed all larvae before the experiment, and experimental larvae were reared in 20 mL of distilled water (DW) in a plastic case with a diameter of 3.5 cm (Figure 1f). The experiment was conducted from June to July 2023. To avoid pollution from food, larvae were not fed during this experiment. The plastic case was covered with a lid to prevent changes in ammonia concentration due to water evaporation.
We reared larvae for 9 days and ammonia concentrations in the rearing water were measured every 3 days (days 0, 3, 6, and 9). We measured the level of dissolved ammonia by Urea/Ammonia Assay Kit (Rapid) (Megazyme, Bray, Ireland) following its protocol, with NanoDrop OneC (Thermo Fisher Scientific, Massachusetts, USA) for wavelength quantification. This assay kit can measure concentrations with 0.018 mg/L as the minimum detectable level. Five microliter samples of rearing water were collected and used for measurements. Since this is a very small amount (0.00025% of total water), the collection of breeding water may not affect the ammonia concentration. In order to account for the body size difference, ammonia concentrations were corrected by individual body weights (concentration [in grams per liter]/body weight [in grams]).
Next, we measured the amount of ammonia in gut. K. eiffingeri was fed with trophic eggs and the other two species were fed with fish food ad libitum, then the feces-containing gut (rectum and large intestine) was taken out from the larva by dissection (Figure 1e). After removing the surface water with paper, we weighed the gut and placed it in microtubes. The gut and its contents were then homogenized and suspended in 100 μL of DW. The ammonia concentration of the suspension was measured using the same protocol as described above. This experiment was performed in August 2023 using K. eiffingeri (n = 10), D. japonicus (n = 10), and R. ornativentris (n = 5).
Larval tolerance to ammonia was experimentally tested using K. eiffingeri (n = 30) and D. japonicus (n = 30) in August 2023. Ammonium chloride is efficient to test the effect of dissolved ammonia on larval survival (Schuytema & Nebeker, 1999). Thirty larvae of each species were individually allocated to three treatments: DW, 50, and 100 mM of ammonium chloride solutions (n = 10 for each, ammonium chloride, FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan). The experimental larvae were checked every 12 h for 7 days for their survival.
Survival rates of each group were compared by Kaplan–Meier curves and log-rank test, followed by Bonferroni correction for multiple comparisons.
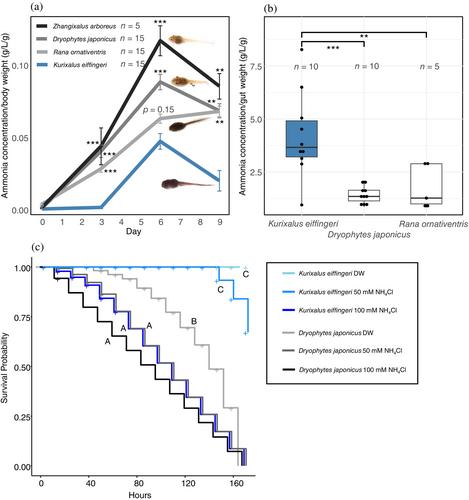
The ammonia concentration of the rearing water was significantly higher except for the comparison with day-6 R. ornativentris (Figure 2a). On the whole, there was a clear pattern that K. eiffingeri produces a very low amount of ammonia in rearing water compared with other species.
The ammonia concentration of the larval gut was significantly higher in K. eiffingeri than in the other two species (Figure 2b, Dunnet's test; p values provided in figure caption) suggesting that K. eiffingeri retain ammonia in their gut during the larval period.
In distilled water, larvae of D. japonicus could not survive more than 7 days (Figure 2c, light gray). In contrast, all larvae of K. eiffingeri survived in distilled water, suggesting that K. eiffingeri is tolerant to the ammonia produced by themselves (Figure 2c, pale light blue). At 50 mM NH4Cl treatment, the majority of K. eiffingeri larvae (8/10, Figure 2c, light blue) survived for 7 days but all D. japonicus larvae died in 7 days (Figure 2c, gray). At the greatest dose of ammonia (100 mM NH4Cl), both K. eiffingeri and D. japonicus larvae could not survive for 7 days (Figure 2c, blue and black), implying that a high dose of ammonia is also toxic to K. eiffingeri.
K. eiffingeri larvae, which grow in closed system tree and stump holes, excrete less ammonia into their environment than other species whose habitats are open water ponds; they also retain high concentrations of ammonia in their gut (Figure 2a,b). This is an indirect indication that K. eiffingeri larvae grow without, or at least reduced, defecation. Additionally, K. eiffingeri larvae were shown to be more tolerant to ammonia than D. japonicus (Figure 2c). These results indicate that the larvae of this species have evolved a unique sanitation strategy to hold the ammonia-containing feces in the larval gut and an unusual tolerance against environmental ammonia. However, K. eiffingeri larvae do emit ammonia, albeit in small amounts, and although the authors have observed that K. eiffingeri larvae do not defecate solid feces, it is possible that they emit ammonia in liquid or other forms, which is an issue that needs further investigation.
For growth and somatic maintenance, animals must take proteinous diets, and consequently, nitrogen excretion is inevitable. Some animals may suffer from the toxic pollution of nitrogen-rich feces. K. eiffingeri firstly defecates just after metamorphosis to adults (Ito, personal observation). This suggests that the nitrogen brought into the phytothermata is eventually carried outside. Females of this species sequentially use the same phytothermata as a breeding site, and as a result, siblings with the same and different clutches grow together and/or sequentially in the same phytothermata (Kam et al., 2000; Ito, personal observation). In this respect, individual sanitation behavior can also work as a social sanitation that eventually raises the fitness of siblings by keeping the growth habitat clean. Such social aspects of sanitation behavior parallel those observed in social insects where larvae of some bees and ants retain their feces in the gut until metamorphosis, bringing the social sanitation benefit to their nestmates (Jackson & Hart, 2009). Growth in narrow, crowding environments with conspecifics, especially siblings, may be a common ecological factor that drives the evolution of the social sanitation behavior.
The question also arises whether similar phenomena can be observed in organisms living in phytotelmata. For example, many species of poison dart frogs (Dendrobatidae) also grow their larvae in phytotelmata (Lima et al., 2007; Summers, 1990; Weygoldt, 1987). We predict that these species may also have similar sanitation strategies. Besides anurans, species that utilize phytotelmata are known in a very wide range of taxa, from protozoa, invertebrates, and vertebrates (Frank, 1983). It will be an important future question to see whether the sanitation adaptation and the tolerances against ammonia are widely observed in various taxonomic groups in this extremely tiny microenvironment. This study has shed light on the novel sanitation behavior and unique biological material flow in phytotelmata, the less conspicuous but frequent and important inland aquatic system for animals (Frank, 1983).
The authors declare no conflicts of interest.
These species are not regulated by the Tokyo Metropolitan University ethics committee or protected by Japanese law. Thus, no ethics approval was required to declare, and no special permits were needed to conduct this research.
期刊介绍:
Ecology publishes articles that report on the basic elements of ecological research. Emphasis is placed on concise, clear articles documenting important ecological phenomena. The journal publishes a broad array of research that includes a rapidly expanding envelope of subject matter, techniques, approaches, and concepts: paleoecology through present-day phenomena; evolutionary, population, physiological, community, and ecosystem ecology, as well as biogeochemistry; inclusive of descriptive, comparative, experimental, mathematical, statistical, and interdisciplinary approaches.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


