Andrew C. Eagar, Princess H. Abu, Megan A. Brown, Sara M. Moledor, Kurt A. Smemo, Richard P. Phillips, Andrea L. Case, Christopher B. Blackwood
{"title":"Setting the stage for plant–soil feedback: Mycorrhizal influences over conspecific recruitment, plant and fungal communities, and coevolution","authors":"Andrew C. Eagar, Princess H. Abu, Megan A. Brown, Sara M. Moledor, Kurt A. Smemo, Richard P. Phillips, Andrea L. Case, Christopher B. Blackwood","doi":"10.1111/1365-2745.14393","DOIUrl":null,"url":null,"abstract":"<h2>1 INTRODUCTION</h2>\n<p>Global environmental change factors are altering plant community composition worldwide (Franklin et al., <span>2016</span>; Komatsu et al., <span>2019</span>) and consequently changing the soil microbial communities that structure plant–soil interactions (Classen et al., <span>2015</span>; Rudgers et al., <span>2020</span>). While the consequences of shifting species' abundances can be hard to predict, aggregated changes in plant communities often result in shifts in the dominance of plant functional types (e.g. mycorrhizal associations). Examples of shifts in mycorrhizal dominance due to changes in plant community composition are now plentiful: ectomycorrhizal (ECM) and ericoid mycorrhizal (ERM) plants are encroaching into grasslands historically dominated by arbuscular mycorrhizal (AM) plants (Brandt et al., <span>2013</span>; Coop & Givnish, <span>2007</span>; Singh, <span>2018</span>); AM plants are expanding into ERM-dominant shrubland ecosystems (Tan et al., <span>2019</span>); ECM plants are invading tundra primarily composed of ERM plants (Elmendorf et al., <span>2012</span>; Myers-Smith et al., <span>2011</span>); and the relative dominance of AM versus ECM trees is changing in temperate forests (Jo et al., <span>2019</span>; Steidinger et al., <span>2019</span>). These shifts in plant–mycorrhizal composition point to a critical need to understand how plant–mycorrhizal types affect plant–soil interactions across plant communities, which have the potential to also alter future ecological and evolutionary dynamics.</p>\n<p>Plants influence soil in ways that have reciprocating positive or negative effects on the growth and reproductive success of themselves and other conspecific individuals, such as their offspring—a process known as ‘plant–soil feedback’ (PSF) (Ehrenfeld et al., <span>2005</span>; van der Putten et al., <span>2013</span>). Although abiotic factors can be important in certain cases (e.g. Xu et al., <span>2021</span>), soil microorganisms are recognized as the primary drivers of PSF (Mangan et al., <span>2010</span>; Mills & Bever, <span>1998</span>; van der Putten et al., <span>2013</span>). Plant pathogens and mutualists, including mycorrhizal fungi, contribute directly to feedback by concomitantly increasing or decreasing in abundance with their associated plant populations. Despite PSF being an outcome of complex interactions with multiple components of the plant microbiome, mycorrhizal type has recently emerged as a particularly important determinant of the net result (positive or negative) of PSF (Bennett et al., <span>2017</span>; Delavaux et al., <span>2023</span>; Eagar et al., <span>2020</span>). Most plant species can be categorized into a single mycorrhizal type (e.g. AM, ECM, or ERM; Brundrett & Tedersoo, <span>2018</span>) and plant communities can be characterized by mycorrhizal type dominance on a continuous scale (e.g. ECM tree basal area/total tree basal area). Thus, plant–mycorrhizal associations represent a powerful conceptual model for generalizing PSF effects and outcomes across species and communities.</p>\n<p>In this review, we discuss how predicted shifts in dominant mycorrhizal types under global change affect plant–soil interactions and related plant community dynamics. We begin by synthesizing the underlying mechanisms and drivers of PSF dynamics among plant species based on their mycorrhizal type, specifically contrasting AM systems to ECM and ERM systems due to trait-based differences that distinguish the former group from the latter two. Next, we explore how PSF generated by dominant species of different mycorrhizal types may ‘spill over’ onto co-occurring plant species, thus affecting the strength and direction of their PSF, and how mycorrhizal spillover may influence plant community dynamics as dominance of a community shifts between mycorrhizal types under global change. This concept is similar to existing plant pathogen spillover theory, where some plants can serve as pathogen reservoirs, enhancing the likelihood of disease in other, nearby plants (Gilbert & Parker, <span>2016</span>; Mordecai, <span>2011</span>; Power & Mitchell, <span>2004</span>). However, the mycorrhizal spillover that we propose here focuses on the effects of dominant plant species on a much broader range of microorganisms (including mutualistic and free-living microorganisms) and the soil biogeochemical environment. We also discuss how mycorrhizal type and related spillover effects can influence plant evolution in communities by affecting drivers of PSF that interact with plant traits related to defence and mutualism formation. We conclude by identifying several topics that warrant further research to fully evaluate these mechanisms and the consequences they may have on plant eco-evolutionary dynamics.</p>","PeriodicalId":191,"journal":{"name":"Journal of Ecology","volume":"16 1","pages":""},"PeriodicalIF":5.3000,"publicationDate":"2024-08-28","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"Journal of Ecology","FirstCategoryId":"93","ListUrlMain":"https://doi.org/10.1111/1365-2745.14393","RegionNum":1,"RegionCategory":"环境科学与生态学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"ECOLOGY","Score":null,"Total":0}
引用次数: 0
Abstract
1 INTRODUCTION
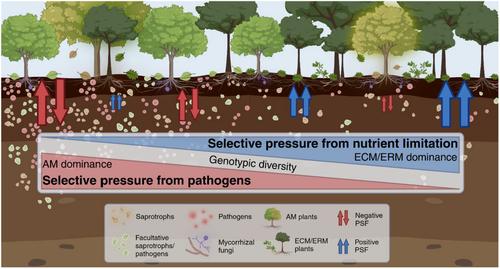
Global environmental change factors are altering plant community composition worldwide (Franklin et al., 2016; Komatsu et al., 2019) and consequently changing the soil microbial communities that structure plant–soil interactions (Classen et al., 2015; Rudgers et al., 2020). While the consequences of shifting species' abundances can be hard to predict, aggregated changes in plant communities often result in shifts in the dominance of plant functional types (e.g. mycorrhizal associations). Examples of shifts in mycorrhizal dominance due to changes in plant community composition are now plentiful: ectomycorrhizal (ECM) and ericoid mycorrhizal (ERM) plants are encroaching into grasslands historically dominated by arbuscular mycorrhizal (AM) plants (Brandt et al., 2013; Coop & Givnish, 2007; Singh, 2018); AM plants are expanding into ERM-dominant shrubland ecosystems (Tan et al., 2019); ECM plants are invading tundra primarily composed of ERM plants (Elmendorf et al., 2012; Myers-Smith et al., 2011); and the relative dominance of AM versus ECM trees is changing in temperate forests (Jo et al., 2019; Steidinger et al., 2019). These shifts in plant–mycorrhizal composition point to a critical need to understand how plant–mycorrhizal types affect plant–soil interactions across plant communities, which have the potential to also alter future ecological and evolutionary dynamics.
Plants influence soil in ways that have reciprocating positive or negative effects on the growth and reproductive success of themselves and other conspecific individuals, such as their offspring—a process known as ‘plant–soil feedback’ (PSF) (Ehrenfeld et al., 2005; van der Putten et al., 2013). Although abiotic factors can be important in certain cases (e.g. Xu et al., 2021), soil microorganisms are recognized as the primary drivers of PSF (Mangan et al., 2010; Mills & Bever, 1998; van der Putten et al., 2013). Plant pathogens and mutualists, including mycorrhizal fungi, contribute directly to feedback by concomitantly increasing or decreasing in abundance with their associated plant populations. Despite PSF being an outcome of complex interactions with multiple components of the plant microbiome, mycorrhizal type has recently emerged as a particularly important determinant of the net result (positive or negative) of PSF (Bennett et al., 2017; Delavaux et al., 2023; Eagar et al., 2020). Most plant species can be categorized into a single mycorrhizal type (e.g. AM, ECM, or ERM; Brundrett & Tedersoo, 2018) and plant communities can be characterized by mycorrhizal type dominance on a continuous scale (e.g. ECM tree basal area/total tree basal area). Thus, plant–mycorrhizal associations represent a powerful conceptual model for generalizing PSF effects and outcomes across species and communities.
In this review, we discuss how predicted shifts in dominant mycorrhizal types under global change affect plant–soil interactions and related plant community dynamics. We begin by synthesizing the underlying mechanisms and drivers of PSF dynamics among plant species based on their mycorrhizal type, specifically contrasting AM systems to ECM and ERM systems due to trait-based differences that distinguish the former group from the latter two. Next, we explore how PSF generated by dominant species of different mycorrhizal types may ‘spill over’ onto co-occurring plant species, thus affecting the strength and direction of their PSF, and how mycorrhizal spillover may influence plant community dynamics as dominance of a community shifts between mycorrhizal types under global change. This concept is similar to existing plant pathogen spillover theory, where some plants can serve as pathogen reservoirs, enhancing the likelihood of disease in other, nearby plants (Gilbert & Parker, 2016; Mordecai, 2011; Power & Mitchell, 2004). However, the mycorrhizal spillover that we propose here focuses on the effects of dominant plant species on a much broader range of microorganisms (including mutualistic and free-living microorganisms) and the soil biogeochemical environment. We also discuss how mycorrhizal type and related spillover effects can influence plant evolution in communities by affecting drivers of PSF that interact with plant traits related to defence and mutualism formation. We conclude by identifying several topics that warrant further research to fully evaluate these mechanisms and the consequences they may have on plant eco-evolutionary dynamics.
期刊介绍:
Journal of Ecology publishes original research papers on all aspects of the ecology of plants (including algae), in both aquatic and terrestrial ecosystems. We do not publish papers concerned solely with cultivated plants and agricultural ecosystems. Studies of plant communities, populations or individual species are accepted, as well as studies of the interactions between plants and animals, fungi or bacteria, providing they focus on the ecology of the plants.
We aim to bring important work using any ecological approach (including molecular techniques) to a wide international audience and therefore only publish papers with strong and ecological messages that advance our understanding of ecological principles.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


