{"title":"A resource for functional investigation of miRNAs in rice responses to viral infection","authors":"Baogang Zhang, Xiong Zhang, Wenji Li, Dezhuo Pan, Baining Ma, Xinhui Duan, Chaoyi Dong, Lu Wang, Mingfu Zhao, Shanshan Zhao, Shuai Zhang, Jianguo Wu","doi":"10.1111/pbi.14455","DOIUrl":null,"url":null,"abstract":"<p>In rice agriculture, the rice grassy stunt virus (RGSV) and rice ragged stunt virus (RRSV) present significant biosafety challenges. RGSV, a <i>Bunyaviridae</i> family virus with single-stranded RNA and RRSV, a double-stranded RNA virus from the Reoviridae family, are mainly transmitted by the brown planthopper, posing a co-infection risk that can cause rice yellowing syndrome. This syndrome can severely hinder plant growth and, in extreme cases, lead to a complete yield loss. RNA interference (RNAi), featuring small RNA molecules such as microRNAs (miRNAs), emerges as a key antiviral defence, modulating gene expression by degrading or inhibiting translation of mRNA. This mechanism is integral to various plant biological processes, including growth, development and stress response. MiR168 and miR528, specifically, have been identified as vital in augmenting rice's resistance to viral attacks (Wu <i>et al</i>., <span>2015</span>, <span>2017</span>). Employing short tandem target mimic (STTM) technology has not only attenuated the function of certain miRNAs in rice but also underscored their role in essential agronomic traits (Zhang <i>et al</i>., <span>2017</span>). Extending this approach, studies in maize and tomato demonstrate miRNAs' influence within hormonal signalling and secondary metabolism (Peng <i>et al</i>., <span>2018</span>). While the regulatory networks of miRNAs in rice are well-documented, the exploration of species-specific miRNAs reactive to viral infections is relatively nascent.</p><p>To advance the understanding of species-specific miRNAs in viral defence, this study established a transgenic rice library covering miRNA overexpression, suppression and gene knockout mutants mediated by CRISPR-Cas9. Small RNA sequencing on rice stem base tissues infected with RGSV or RRSV identified 23 miRNAs with at least a two-fold expression change relative to uninfected controls (Table S1). Heatmap analysis highlighted differential miRNA response patterns to RGSV and RRSV infections (Figure 1a). Validation by qRT-PCR corroborated the altered expression of six <i>miRNAs</i> (<i>MIR440</i>, <i>MIR535</i>, <i>MIR1846d</i>, <i>MIR1863a</i>, <i>MIR1874-3p</i> and <i>MIR1881</i>), consistent with sequencing results. Notably, miR440, miR1846d, miR1863a and miR1874-3p were predominantly downregulated by RGSV, whereas miR535 and miR1881 were suppressed by both viruses (Figure 1b,c). To elucidate miRNA roles in viral pathogenesis, we engineered three vector types for precise miRNA regulation. The overexpression vector was constructed by cloning the miRNA precursor with ~100 bp flanking sequences downstream of the Actin1 promoter (Figure S1a). For knockdown, the STTM technology was employed; we fused the miRNA complementary sequence plus three nucleotides to a 48 nt linker, and inserted this downstream of the Actin1 promoter to create the STTM vector (Figure S1b). Additionally, we designed a CRISPR-Cas9 editing vector by targeting two sites on the miRNA precursor sequence (Figure S1c). We successfully introduced 68 vectors into the ZH11 rice variety using Agrobacterium-mediated transformation, achieving 20 overexpression, 22 STTM-mediated knockdown and 20 CRISPR-Cas9-mediated knockout events. Each event type was confirmed in at least two independent transgenic lines, fulfilling the expected experimental criteria (Figures S2–S4; Tables S2 and S3). The phenotypic profiles of select transgenic lines at the booting stage are documented in Figure S5. In T2 generation homozygous lines, we performed inoculation trials with RGSV and RRSV to determine the miRNAs' roles in rice-virus interactions. Post-RGSV inoculation, the overexpression line <i>OX535</i> demonstrated a significant susceptibility rate of 76.9%, while the <i>MIM535</i> suppression line and the <i>mir535</i> knockout line showed enhanced resistance (Table S4). <i>MIR535</i> is ubiquitously expressed in rice tissues, including rhizomes, stem base, glumes and germinated seeds (Figure S6). Furthermore, RGSV infection was found to downregulate <i>MIR535</i> promoter activity, reducing <i>MIR535</i> transcript levels (Figures 1b,d and S7). Pathogenic assessments and virus titre analyses revealed that the <i>OX535</i> line accumulated greater viral loads and exhibited more severe symptoms, such as pronounced dwarfing and increased tillering. In contrast, the <i>MIM535</i> suppression line and the <i>mir535</i> knockout line presented with lower virus titre and milder symptoms (Figure 1e–g). Under non-stress conditions, <i>OX535</i> displayed dwarfism, multiple tillering, smaller spikes and reduced spikelet branching, but grains were longer and wider. No significant variances were observed in the <i>MIM535</i> and <i>mir535</i> lines for key agronomic traits, including plant height, tiller number, spike size, grain dimensions and 100-grain weight, compared to the ZH11 variety (Figures 1h–n and S8). The enhanced viral resistance of these lines identifies miR535 as a promising target for RGSV resistance breeding.</p><p>Rice ragged stunt virus resistance screening indicated that the <i>OX1868.1</i> line exhibited enhanced viral resistance, while the <i>MIM1868.1</i> line was more susceptible to the virus. Multiple lines of evidence indicate that miR1868.1 can suppress RRSV accumulation. This includes quantifying viral transcript levels using RT-qPCR, and detecting the expression levels of viral coat proteins through Western blot analysis (Figure 1o–q). Agronomic trait assessments showed no significant differences in panicle length, grain size and 100-grain weight between <i>OX1868.1</i>, <i>MIM1868.1</i> and the ZH11 control, although <i>OX1868.1</i> exhibited reduced tillering and decreased plant height (Figures 1r–x and S9). Consequently, miR1868.1 demonstrates antiviral activity against RRSV and represents a valuable target for rice antiviral breeding programs.</p><p>The research has successfully established a miRNA resource library with modalities for overexpression, mimicry (MIM) and knockout of miRNAs. This library facilitates the modelling of gene loss-of-function and the resulting phenotypic manifestations. Of the 23 miRNAs curated, 16 exist as single copies in rice, potentially increasing the effectiveness of CRISPR-Cas9 knockouts. Remarkably, 19 miRNAs are so far exclusive to rice, indicating a library enriched with species-specific sequences. Beyond the previously mentioned miR535 and miR1868.1, this work extends to other miRNAs involved in the viral resistance of rice. The genetic constructs are also valuable for investigating rice growth, development and response to biotic and abiotic stress. Overall, this initiative has created a transgenic rice miRNA library, spotlighting 23 miRNAs pivotal to viral infection responses.</p><p>The authors have declared no conflict of interest.</p><p>B.Z., S.-S.Z. and J.W. designed the experiments. B.Z., X.Z., W.L., D.P., B.M., X.D., C.D., L.W. and M.Z. conducted the experiments and analysed the data. B.Z. and S.Z. wrote the paper with the input of all other authors.</p>","PeriodicalId":221,"journal":{"name":"Plant Biotechnology Journal","volume":"22 12","pages":"3380-3382"},"PeriodicalIF":10.1000,"publicationDate":"2024-08-24","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1111/pbi.14455","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"Plant Biotechnology Journal","FirstCategoryId":"5","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1111/pbi.14455","RegionNum":1,"RegionCategory":"生物学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"BIOTECHNOLOGY & APPLIED MICROBIOLOGY","Score":null,"Total":0}
引用次数: 0
Abstract
In rice agriculture, the rice grassy stunt virus (RGSV) and rice ragged stunt virus (RRSV) present significant biosafety challenges. RGSV, a Bunyaviridae family virus with single-stranded RNA and RRSV, a double-stranded RNA virus from the Reoviridae family, are mainly transmitted by the brown planthopper, posing a co-infection risk that can cause rice yellowing syndrome. This syndrome can severely hinder plant growth and, in extreme cases, lead to a complete yield loss. RNA interference (RNAi), featuring small RNA molecules such as microRNAs (miRNAs), emerges as a key antiviral defence, modulating gene expression by degrading or inhibiting translation of mRNA. This mechanism is integral to various plant biological processes, including growth, development and stress response. MiR168 and miR528, specifically, have been identified as vital in augmenting rice's resistance to viral attacks (Wu et al., 2015, 2017). Employing short tandem target mimic (STTM) technology has not only attenuated the function of certain miRNAs in rice but also underscored their role in essential agronomic traits (Zhang et al., 2017). Extending this approach, studies in maize and tomato demonstrate miRNAs' influence within hormonal signalling and secondary metabolism (Peng et al., 2018). While the regulatory networks of miRNAs in rice are well-documented, the exploration of species-specific miRNAs reactive to viral infections is relatively nascent.
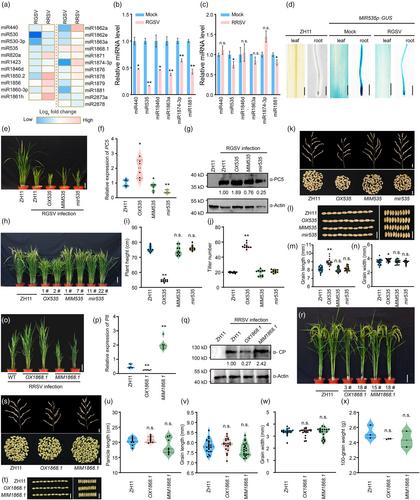
To advance the understanding of species-specific miRNAs in viral defence, this study established a transgenic rice library covering miRNA overexpression, suppression and gene knockout mutants mediated by CRISPR-Cas9. Small RNA sequencing on rice stem base tissues infected with RGSV or RRSV identified 23 miRNAs with at least a two-fold expression change relative to uninfected controls (Table S1). Heatmap analysis highlighted differential miRNA response patterns to RGSV and RRSV infections (Figure 1a). Validation by qRT-PCR corroborated the altered expression of six miRNAs (MIR440, MIR535, MIR1846d, MIR1863a, MIR1874-3p and MIR1881), consistent with sequencing results. Notably, miR440, miR1846d, miR1863a and miR1874-3p were predominantly downregulated by RGSV, whereas miR535 and miR1881 were suppressed by both viruses (Figure 1b,c). To elucidate miRNA roles in viral pathogenesis, we engineered three vector types for precise miRNA regulation. The overexpression vector was constructed by cloning the miRNA precursor with ~100 bp flanking sequences downstream of the Actin1 promoter (Figure S1a). For knockdown, the STTM technology was employed; we fused the miRNA complementary sequence plus three nucleotides to a 48 nt linker, and inserted this downstream of the Actin1 promoter to create the STTM vector (Figure S1b). Additionally, we designed a CRISPR-Cas9 editing vector by targeting two sites on the miRNA precursor sequence (Figure S1c). We successfully introduced 68 vectors into the ZH11 rice variety using Agrobacterium-mediated transformation, achieving 20 overexpression, 22 STTM-mediated knockdown and 20 CRISPR-Cas9-mediated knockout events. Each event type was confirmed in at least two independent transgenic lines, fulfilling the expected experimental criteria (Figures S2–S4; Tables S2 and S3). The phenotypic profiles of select transgenic lines at the booting stage are documented in Figure S5. In T2 generation homozygous lines, we performed inoculation trials with RGSV and RRSV to determine the miRNAs' roles in rice-virus interactions. Post-RGSV inoculation, the overexpression line OX535 demonstrated a significant susceptibility rate of 76.9%, while the MIM535 suppression line and the mir535 knockout line showed enhanced resistance (Table S4). MIR535 is ubiquitously expressed in rice tissues, including rhizomes, stem base, glumes and germinated seeds (Figure S6). Furthermore, RGSV infection was found to downregulate MIR535 promoter activity, reducing MIR535 transcript levels (Figures 1b,d and S7). Pathogenic assessments and virus titre analyses revealed that the OX535 line accumulated greater viral loads and exhibited more severe symptoms, such as pronounced dwarfing and increased tillering. In contrast, the MIM535 suppression line and the mir535 knockout line presented with lower virus titre and milder symptoms (Figure 1e–g). Under non-stress conditions, OX535 displayed dwarfism, multiple tillering, smaller spikes and reduced spikelet branching, but grains were longer and wider. No significant variances were observed in the MIM535 and mir535 lines for key agronomic traits, including plant height, tiller number, spike size, grain dimensions and 100-grain weight, compared to the ZH11 variety (Figures 1h–n and S8). The enhanced viral resistance of these lines identifies miR535 as a promising target for RGSV resistance breeding.
Rice ragged stunt virus resistance screening indicated that the OX1868.1 line exhibited enhanced viral resistance, while the MIM1868.1 line was more susceptible to the virus. Multiple lines of evidence indicate that miR1868.1 can suppress RRSV accumulation. This includes quantifying viral transcript levels using RT-qPCR, and detecting the expression levels of viral coat proteins through Western blot analysis (Figure 1o–q). Agronomic trait assessments showed no significant differences in panicle length, grain size and 100-grain weight between OX1868.1, MIM1868.1 and the ZH11 control, although OX1868.1 exhibited reduced tillering and decreased plant height (Figures 1r–x and S9). Consequently, miR1868.1 demonstrates antiviral activity against RRSV and represents a valuable target for rice antiviral breeding programs.
The research has successfully established a miRNA resource library with modalities for overexpression, mimicry (MIM) and knockout of miRNAs. This library facilitates the modelling of gene loss-of-function and the resulting phenotypic manifestations. Of the 23 miRNAs curated, 16 exist as single copies in rice, potentially increasing the effectiveness of CRISPR-Cas9 knockouts. Remarkably, 19 miRNAs are so far exclusive to rice, indicating a library enriched with species-specific sequences. Beyond the previously mentioned miR535 and miR1868.1, this work extends to other miRNAs involved in the viral resistance of rice. The genetic constructs are also valuable for investigating rice growth, development and response to biotic and abiotic stress. Overall, this initiative has created a transgenic rice miRNA library, spotlighting 23 miRNAs pivotal to viral infection responses.
The authors have declared no conflict of interest.
B.Z., S.-S.Z. and J.W. designed the experiments. B.Z., X.Z., W.L., D.P., B.M., X.D., C.D., L.W. and M.Z. conducted the experiments and analysed the data. B.Z. and S.Z. wrote the paper with the input of all other authors.
期刊介绍:
Plant Biotechnology Journal aspires to publish original research and insightful reviews of high impact, authored by prominent researchers in applied plant science. The journal places a special emphasis on molecular plant sciences and their practical applications through plant biotechnology. Our goal is to establish a platform for showcasing significant advances in the field, encompassing curiosity-driven studies with potential applications, strategic research in plant biotechnology, scientific analysis of crucial issues for the beneficial utilization of plant sciences, and assessments of the performance of plant biotechnology products in practical applications.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


