{"title":"Ustilaginoidea virens secreted effector UvSec117 hijacks OsWRKY31-OsAOC module to suppress jasmonic acid-mediated immunity in rice","authors":"Yuhang Duan, Guogen Yang, Jintian Tang, Yuan Fang, Hailin Wang, Zhaoyun Wang, Hao Liu, Xiaolin Chen, Junbin Huang, Jing Chen, Qiutao Xu, Lu Zheng, Xiaoyang Chen","doi":"10.1111/pbi.14452","DOIUrl":null,"url":null,"abstract":"<p>Rice false smut (RFS) caused by <i>Ustilaginoidea virens</i> is one of the most important disease in rice (<i>Oryza sativa</i>)-growing regions worldwide. RFS not only causes rice yield losses but also potentially threatens human and animal health by producing cyclopeptide mycotoxins (Sun <i>et al</i>., <span>2020</span>). Introducing genetically encoded resistance is an environmentally friendly, economical approach to controlling plant diseases (Yu <i>et al</i>., <span>2023</span>). However, at present, the varieties and gene resources of resistance to RFS are still extremely scarce, and it is difficult to identify major resistance genes against RFS. Uncovering the functions of the <i>U. virens</i> effectors and molecular mechanism of the rice, <i>U. virens</i> interaction can help to identify molecular probes for discovering disease resistance-related genes (Wang and Kawano, <span>2022</span>).</p><p>In previous studies, we identified UvSec117 as a key virulence effector in <i>U. virens</i>, and found rice transcription factor OsWRKY31 in a screen for proteins that interact with UvSec117 (Chen <i>et al</i>., <span>2022</span>). WRKY transcription factors have many regulatory roles in development and response to biotic/abiotic stresses in plants (Wang <i>et al</i>., <span>2023</span>). However, little is known about the regulatory functions of WRKY genes in the plant resistance to grain-infecting pathogens. In this work, we confirmed interactions between UvSec117 and OsWRKY31 in a directed yeast two-hybrid assay (Figure 1a; Data S1). In a co-immunoprecipitation (Co-IP) assay by rice protoplasts transiently co-expressing <i>OsWRKY31-Flag</i> and <i>UvSec117-GFP</i> constructs, UvSec117 was immunoprecipitated by OsWRKY31 (Figure 1b). In a pull-down assay using recombinant OsWRKY31-GST and UvSec117-His purified from <i>Escherichia coli</i>, OsWRKY31-GST was pulled down by His beads coated with UvSec117-His (Figure 1c). We also validated the interaction between UvSec117 and OsWRKY31 by a luciferase complementation imaging (LCI) assay in <i>N. benthamiana</i> leaves (Figure 1d). When we transiently co-expressed <i>UvSec117-cYFP</i> and <i>OsWRKY31-nYFP</i> constructs in rice protoplasts and performed a bimolecular fluorescence complementation (BiFC) assay, we detected YFP (yellow fluorescent protein) fluorescence in the nucleus (Figure 1e). Collectively, these results suggest that UvSec117 interacts with OsWRKY31 in vivo and in vitro.</p><p>To explore the role of OsWRKY31 in resistance against RFS fungus or other rice pathogens, we generated <i>OsWRKY31</i> knockout mutant plants (<i>wrky31</i>) (Figure S1a) and <i>OsWRKY31</i>-overexpressing transgenic rice lines (<i>OsWRKY31</i>-OE) (Figure S1b). The agronomic traits of <i>wrky31</i> and <i>OsWRKY31</i>-OE plants were similar to those of wild-type <i>Nipponbare</i> (NPB) (Figure S1c,d). Following inoculation with different rice pathogens, <i>OsWRKY31</i>-OE plants were less susceptible and <i>wrky31</i> plants were more susceptible to the RFS, bacterial blight, rice blast and sheath blight than NPB plants (Figure 1f–i), indicating that OsWRKY31 positively regulates the resistance of rice to multiple diseases.</p><p>To identify global targets of the transcription factor OsWRKY31, we performed chromatin immunoprecipitation followed by deep sequencing (ChIP-seq) using <i>OsWRKY31</i>-OE plants with an anti-Flag antibody. In total, we identified 4626 peaks (1054 target genes, Data S2). A significant majority (> 60%) of these peaks are located within genic regions, with the modifications being highly enriched at the promoters of protein-coding genes (Figure 1j). MEME (Multiple EM for Motif Elicitation) analysis revealed that most OsWRKY31-bound DNA motifs contained the sequence TTGTACTT, GGGCCCAC or CCCCTTTT (Figure 1k). Gene ontology (GO) analysis revealed that the target genes were enriched for induced systemic resistance and salicylic acid (SA)/jasmonic acid (JA)-mediated signalling pathways (Figure 1l). RT-qPCR showed that the key JA biosynthesis gene <i>OsAOC</i> (<i>ALLENE OXIDE CYCLASE</i>) is significantly downregulated in <i>wrky31</i>-1 plants, and ChIP-qPCR confirmed that OsWRKY31 binds to the <i>OsAOC</i> promoter (Figure 1m). Knockout of <i>OsAOC</i> in rice enhances its susceptibility to RFS (Figure 1n). <i>OsWRKY31</i> expression in <i>N. benthamiana</i> significantly enhanced firefly luciferase (LUC) activity derived from the <i>OsAOCpro</i>-<i>LUC</i> reporter. Co-infiltration of <i>UvSec117</i> with <i>OsAOCpro</i>-<i>LUC</i> inhibited OsWRKY31-induced LUC activity, whereas co-infiltration of <i>GFP</i>, did not (Figure 1o). Yeast one-hybrid results showed that OsWRKY31 can bind the promoter of <i>OsAOC</i> (Figure 1p). In an electrophoretic mobility shift assay (EMSA) assay, OsWRKY31-His specifically bound to the <i>OsAOC</i> promoter; addition of unlabelled competitive probe decreased this binding. Preincubation with UvSec117 reduced the DNA-binding activity of OsWRKY31 (Figure 1q), indicating that UvSec117 directly inhibits the DNA-binding activity of OsWRKY31. Moreover, the contents of JA were significantly lower in <i>wrky31</i>-1 than in NPB rice spikelets; in HE-1 (Heterologous expression of <i>UvSec117</i> transgenic plants) relative to EV (empty vector transgenic plants) rice spikelets (Figure 1r). These results indicate that OsWRKY31 regulating the JA-mediated defence was suppressed by UvSec117.</p><p>In this study, we found that the transcription factor OsWRKY31 functions as a key positive regulator to broad-spectrum disease resistance. Here, we provide a comprehensive genome-wide binding map of OsWRKY31 and its regulatory network, and further describe a previously unknown regulatory role where OsWRKY31 mediates the JA-mediated signalling pathway to regulate plant immunity. Collectively, this study unveils a pivotal virulence strategy employed by <i>U. virens</i>, the secretory effector UvSec117 inhibits OsWRKY31 binding to target gene promoters like <i>OsAOC</i>, thereby suppressing JA-mediated defence (Figure 1s). Moreover, this investigation highlights the critical role of OsWRKY31 as a crucial component in orchestrating multi-pathogen resistance, further underscoring its significance in plant defence mechanisms. The <i>OsWRKY31</i>-OE lines generated in this study may provide valuable germplasm resources for rice disease resistance breeding, which has important theoretical and practical value.</p><p>The authors declare no conflict of interest.</p><p>Y.D. J.T. and G.Y. performed most of the experiments. Q.X. performed the data analyses. L.Z., H.L., H.W., Z.W., Y.F., J.H., J.C. and X-L.C. provided technical support. X-Y.C. and L.Z. wrote and revised the manuscript. All authors have read and approved the final manuscript.</p>","PeriodicalId":221,"journal":{"name":"Plant Biotechnology Journal","volume":"22 12","pages":"3342-3344"},"PeriodicalIF":10.5000,"publicationDate":"2024-08-16","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1111/pbi.14452","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"Plant Biotechnology Journal","FirstCategoryId":"5","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1111/pbi.14452","RegionNum":1,"RegionCategory":"生物学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"BIOTECHNOLOGY & APPLIED MICROBIOLOGY","Score":null,"Total":0}
引用次数: 0
Abstract
Rice false smut (RFS) caused by Ustilaginoidea virens is one of the most important disease in rice (Oryza sativa)-growing regions worldwide. RFS not only causes rice yield losses but also potentially threatens human and animal health by producing cyclopeptide mycotoxins (Sun et al., 2020). Introducing genetically encoded resistance is an environmentally friendly, economical approach to controlling plant diseases (Yu et al., 2023). However, at present, the varieties and gene resources of resistance to RFS are still extremely scarce, and it is difficult to identify major resistance genes against RFS. Uncovering the functions of the U. virens effectors and molecular mechanism of the rice, U. virens interaction can help to identify molecular probes for discovering disease resistance-related genes (Wang and Kawano, 2022).
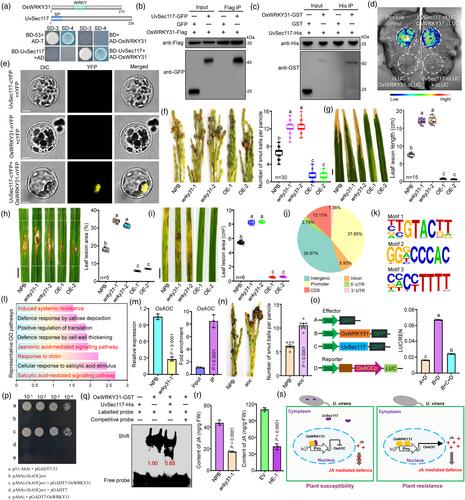
In previous studies, we identified UvSec117 as a key virulence effector in U. virens, and found rice transcription factor OsWRKY31 in a screen for proteins that interact with UvSec117 (Chen et al., 2022). WRKY transcription factors have many regulatory roles in development and response to biotic/abiotic stresses in plants (Wang et al., 2023). However, little is known about the regulatory functions of WRKY genes in the plant resistance to grain-infecting pathogens. In this work, we confirmed interactions between UvSec117 and OsWRKY31 in a directed yeast two-hybrid assay (Figure 1a; Data S1). In a co-immunoprecipitation (Co-IP) assay by rice protoplasts transiently co-expressing OsWRKY31-Flag and UvSec117-GFP constructs, UvSec117 was immunoprecipitated by OsWRKY31 (Figure 1b). In a pull-down assay using recombinant OsWRKY31-GST and UvSec117-His purified from Escherichia coli, OsWRKY31-GST was pulled down by His beads coated with UvSec117-His (Figure 1c). We also validated the interaction between UvSec117 and OsWRKY31 by a luciferase complementation imaging (LCI) assay in N. benthamiana leaves (Figure 1d). When we transiently co-expressed UvSec117-cYFP and OsWRKY31-nYFP constructs in rice protoplasts and performed a bimolecular fluorescence complementation (BiFC) assay, we detected YFP (yellow fluorescent protein) fluorescence in the nucleus (Figure 1e). Collectively, these results suggest that UvSec117 interacts with OsWRKY31 in vivo and in vitro.
To explore the role of OsWRKY31 in resistance against RFS fungus or other rice pathogens, we generated OsWRKY31 knockout mutant plants (wrky31) (Figure S1a) and OsWRKY31-overexpressing transgenic rice lines (OsWRKY31-OE) (Figure S1b). The agronomic traits of wrky31 and OsWRKY31-OE plants were similar to those of wild-type Nipponbare (NPB) (Figure S1c,d). Following inoculation with different rice pathogens, OsWRKY31-OE plants were less susceptible and wrky31 plants were more susceptible to the RFS, bacterial blight, rice blast and sheath blight than NPB plants (Figure 1f–i), indicating that OsWRKY31 positively regulates the resistance of rice to multiple diseases.
To identify global targets of the transcription factor OsWRKY31, we performed chromatin immunoprecipitation followed by deep sequencing (ChIP-seq) using OsWRKY31-OE plants with an anti-Flag antibody. In total, we identified 4626 peaks (1054 target genes, Data S2). A significant majority (> 60%) of these peaks are located within genic regions, with the modifications being highly enriched at the promoters of protein-coding genes (Figure 1j). MEME (Multiple EM for Motif Elicitation) analysis revealed that most OsWRKY31-bound DNA motifs contained the sequence TTGTACTT, GGGCCCAC or CCCCTTTT (Figure 1k). Gene ontology (GO) analysis revealed that the target genes were enriched for induced systemic resistance and salicylic acid (SA)/jasmonic acid (JA)-mediated signalling pathways (Figure 1l). RT-qPCR showed that the key JA biosynthesis gene OsAOC (ALLENE OXIDE CYCLASE) is significantly downregulated in wrky31-1 plants, and ChIP-qPCR confirmed that OsWRKY31 binds to the OsAOC promoter (Figure 1m). Knockout of OsAOC in rice enhances its susceptibility to RFS (Figure 1n). OsWRKY31 expression in N. benthamiana significantly enhanced firefly luciferase (LUC) activity derived from the OsAOCpro-LUC reporter. Co-infiltration of UvSec117 with OsAOCpro-LUC inhibited OsWRKY31-induced LUC activity, whereas co-infiltration of GFP, did not (Figure 1o). Yeast one-hybrid results showed that OsWRKY31 can bind the promoter of OsAOC (Figure 1p). In an electrophoretic mobility shift assay (EMSA) assay, OsWRKY31-His specifically bound to the OsAOC promoter; addition of unlabelled competitive probe decreased this binding. Preincubation with UvSec117 reduced the DNA-binding activity of OsWRKY31 (Figure 1q), indicating that UvSec117 directly inhibits the DNA-binding activity of OsWRKY31. Moreover, the contents of JA were significantly lower in wrky31-1 than in NPB rice spikelets; in HE-1 (Heterologous expression of UvSec117 transgenic plants) relative to EV (empty vector transgenic plants) rice spikelets (Figure 1r). These results indicate that OsWRKY31 regulating the JA-mediated defence was suppressed by UvSec117.
In this study, we found that the transcription factor OsWRKY31 functions as a key positive regulator to broad-spectrum disease resistance. Here, we provide a comprehensive genome-wide binding map of OsWRKY31 and its regulatory network, and further describe a previously unknown regulatory role where OsWRKY31 mediates the JA-mediated signalling pathway to regulate plant immunity. Collectively, this study unveils a pivotal virulence strategy employed by U. virens, the secretory effector UvSec117 inhibits OsWRKY31 binding to target gene promoters like OsAOC, thereby suppressing JA-mediated defence (Figure 1s). Moreover, this investigation highlights the critical role of OsWRKY31 as a crucial component in orchestrating multi-pathogen resistance, further underscoring its significance in plant defence mechanisms. The OsWRKY31-OE lines generated in this study may provide valuable germplasm resources for rice disease resistance breeding, which has important theoretical and practical value.
The authors declare no conflict of interest.
Y.D. J.T. and G.Y. performed most of the experiments. Q.X. performed the data analyses. L.Z., H.L., H.W., Z.W., Y.F., J.H., J.C. and X-L.C. provided technical support. X-Y.C. and L.Z. wrote and revised the manuscript. All authors have read and approved the final manuscript.
期刊介绍:
Plant Biotechnology Journal aspires to publish original research and insightful reviews of high impact, authored by prominent researchers in applied plant science. The journal places a special emphasis on molecular plant sciences and their practical applications through plant biotechnology. Our goal is to establish a platform for showcasing significant advances in the field, encompassing curiosity-driven studies with potential applications, strategic research in plant biotechnology, scientific analysis of crucial issues for the beneficial utilization of plant sciences, and assessments of the performance of plant biotechnology products in practical applications.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


