{"title":"A new START","authors":"Michael Palmgren, Rosa Laura López-Marqués","doi":"10.1111/nph.20010","DOIUrl":null,"url":null,"abstract":"<p>In higher plants, the outermost cell layer of the apical meristem gives rise to the epidermis of the primary shoot. PDF2 and its paralog ARABIDOPSIS THALIANA MERISTEM LAYER 1 (ATML1) are class IV homeodomain leucine-zipper (HD-Zip IV) transcription factors that are redundantly required for specification of the epidermis (Abe <i>et al</i>., <span>2003</span>; Schrick <i>et al</i>., <span>2023</span>). These transcription factors are activated by gibberellic acid, which leads to the activation of gene expression and downstream events involved in the elongation of epidermal cells and the coordination of epidermal growth with inner tissues (Rombolá-Caldentey <i>et al</i>., <span>2014</span>).</p><p>A common feature of HD-Zip IV transcription factors, including PDF2 and ATML1, is the presence of a lipid sensor, the steroidogenic acute regulatory protein (StAR)-related lipid transfer (START) domain, which forms a hydrophobic pocket predicted to bind lipid partners (Schrick <i>et al</i>., <span>2004</span>). START domain-containing proteins were first characterized in mammals, where they act in lipid signaling and transport (Ponting & Aravind, <span>1999</span>), but their presence in transcription factors is unique to plants (Schrick <i>et al</i>., <span>2004</span>).</p><p>To investigate what kind of ligand(s) binds to the START domain of PDF2, Arabidopsis cell lines were generated by overexpressing either full-length PDF2 or mutants lacking the START domain. The recombinant proteins were tagged with a tandem affinity purification (TAP)-tag and TAP-tagged proteins were immunoprecipitated from cell lysates. After using mass-spectrometry (MS)-based proteomics to confirm the purification of PDF2, a liquid-chromatography MS (LC-MS) platform that targeted more than 240 lipid species was used to identify lipids co-purifying with PDF2. This analysis revealed that 12 lipid species were at least four-fold enriched in samples containing PDF2 compared to the mutant protein lacking the START domain. The lipid species with the highest degree of enrichment (seven to 12-fold) was lyso-PC.</p><p>To confirm that lyso-PC binds to the START domain, a fusion between the START domain and maltose binding protein (MBP) was expressed in <i>Escherichia coli</i>. As a control, a similar fusion protein mutated in the presumed lipid-binding pocket of the START domain was also expressed. After MBP cleavage, the interaction between the START domains and lyso-PC was studied by analyzing changes in fluorescence in microscale thermophoresis (MST) assays. In this assay, both the wild-type (WT) START domain and the mutant form bound phosphatidylcholine (PC) liposomes with the same affinity. However, in a reconstituted system where liposomes were doped with lyso-PC, the WT START domain bound the liposomes with an affinity <i>c</i>. 12-fold higher than that of the mutant.</p><p>In summary, purification of PDF2 from Arabidopsis cell cultures revealed enrichment in lyso-PCs, and direct interaction between the PDF2 START domain and lyso-PCs was verified by <i>in vitro</i> binding assays using liposomes. This suggested to the authors that PDF2 functions as a lyso-PC sensor via its START domain.</p><p>DNA-PDF2 electrophoretic mobility shift assays (EMSA) further demonstrated that PDF2 binds an octameric DNA promoter motif implicated in phospholipid remodeling and the phosphate starvation response (Wojciechowska <i>et al</i>.). Perfect matches were found in the promoters or 5′-UTR regions of glycerophosphodiester phosphodiesterase (<i>GDPD1</i>), the SPX domain-containing nuclear protein <i>SPX1</i>, and a nonspecific phospholipase C (<i>NPC4</i>), amongst other genes. Subsequent quantitative reverse transcription polymerase chain reaction analysis demonstrated that PDF2 is a transcriptional repressor for <i>GDPD1</i> and <i>SPX1</i>. While a clear repression or activation pattern was not found for <i>NPC4</i> in response to PDF2, this gene is highly upregulated during phosphate starvation and PDF2 might contribute to the modulation of this response (Yang <i>et al</i>., <span>2021</span>; Wojciechowska <i>et al</i>.). Together with the results of lipid-binding assays, this suggests that Arabidopsis PDF2 can control gene expression by sensing lipids in an analogous way as nuclear receptors operate in animals.</p><p>Root length was found to be reduced in <i>pdf2</i> mutants and enlarged in <i>PDF2</i> overexpressors, which suggested that PDF2 is important for elongation growth. As a possible mechanism, it is suggested that PDF2 transcriptionally regulates the synthesis of phospholipids required for membrane elongation. It is well known that phosphate starvation triggers a response where primary root growth is arrested and compensated by lateral root formation and root hair growth (Péret <i>et al</i>., <span>2011</span>). Thus, by sensing the phospholipid status in the epidermis when inorganic phosphate is limited, the activity of PDF2 may adjust the synthesis of membrane lipids to trigger a root growth response (Wojciechowska <i>et al</i>., <span>2024</span>).</p><p>What could be the physiological implications of lyso-PCs binding to PDF2 (Fig. 1)? Under phosphate starvation, different phospholipases are activated to trigger the replacement of phospholipids by galactolipids and/or the accumulation of triacylglycerols (TAGs), which allows the plant cells to recycle phosphate (Roychowdhury <i>et al</i>., <span>2023</span>). This ultimately results in a marked decrease in lyso-PC levels, but transient increases in lysolipid levels have been documented for other plant responses requiring membrane remodeling (Roychowdhury <i>et al</i>., <span>2023</span>). In addition, extracellular lyso-PC is actively taken up into the roots of Arabidopsis by the lipid flippase ALA10, which might allow the plant to sense the availability of inorganic phosphate from decaying humus in the soil (Poulsen <i>et al</i>., <span>2015</span>). It has long been known that lyso-PCs activate the plasma membrane H<sup>+</sup>-ATPase (Palmgren & Sommarin, <span>1989</span>), which provides the driving force for uptake of phosphate in root epidermal cells, and induction of phosphate transporters in response to lyso-PC levels has been documented in arbuscular mycorrhizal symbioses in tomato and during Zn starvation responses in Arabidopsis (Drissner <i>et al</i>., <span>2007</span>; Kisko <i>et al</i>., <span>2018</span>). Thus, the results from Wojciechowska <i>et al</i>. suggest that fast changes in lyso-PC levels due to PC degradation and flippase internalization may activate not only the plasma membrane H<sup>+</sup>-ATPase, but also the phosphate starvation response via PDF2. In this way, PDF2 provides a link between the metabolic states and cell growth and differentiation.</p><p>An open question in the work of Wojciechowska <i>et al</i>. is how lyso-PC and PDF2 meet in the cell. While no answer to this question is provided, Wojciechowska <i>et al</i>. suggest that a co-repressor acts with PDF2. One possibility is that the co-repressor binds lyso-PC before travelling to the nucleus and delivers the lipid to PDF2. Alternatively, cytosolic PDF2 would directly bind lyso-PC and then travel to the nucleus to exert its function.</p><p>Last but not least, lyso-PC is proposed to be a signaling lipid, not only during phosphate starvation, but also in other environmental stresses (cold, heat, pathogens) (Hou <i>et al</i>., <span>2016</span>). It has been suggested that the phosphate starvation response overlaps with the responses to these other stresses by use of shared transcription factors (Baek <i>et al</i>., <span>2017</span>). Thus, the work of Wojciechowska <i>et al</i>. generates several questions. What other plant HD-Zip IV transcription factors sense lyso-PC? Is PDF2 involved in lyso-PC-triggered responses to other stresses? Several lipid species different from lyso-PC were identified in lipidomics analysis of full-length PDF2 purified from plant cells. Can the START domain of PDF2 bind different lipids with different affinities? Is this used as a mechanism to sense different types of stresses that might result in distinctly altered lipid ratios?</p>","PeriodicalId":214,"journal":{"name":"New Phytologist","volume":null,"pages":null},"PeriodicalIF":8.3000,"publicationDate":"2024-07-30","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1111/nph.20010","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"New Phytologist","FirstCategoryId":"99","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1111/nph.20010","RegionNum":1,"RegionCategory":"生物学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"PLANT SCIENCES","Score":null,"Total":0}
引用次数: 0
Abstract
In higher plants, the outermost cell layer of the apical meristem gives rise to the epidermis of the primary shoot. PDF2 and its paralog ARABIDOPSIS THALIANA MERISTEM LAYER 1 (ATML1) are class IV homeodomain leucine-zipper (HD-Zip IV) transcription factors that are redundantly required for specification of the epidermis (Abe et al., 2003; Schrick et al., 2023). These transcription factors are activated by gibberellic acid, which leads to the activation of gene expression and downstream events involved in the elongation of epidermal cells and the coordination of epidermal growth with inner tissues (Rombolá-Caldentey et al., 2014).
A common feature of HD-Zip IV transcription factors, including PDF2 and ATML1, is the presence of a lipid sensor, the steroidogenic acute regulatory protein (StAR)-related lipid transfer (START) domain, which forms a hydrophobic pocket predicted to bind lipid partners (Schrick et al., 2004). START domain-containing proteins were first characterized in mammals, where they act in lipid signaling and transport (Ponting & Aravind, 1999), but their presence in transcription factors is unique to plants (Schrick et al., 2004).
To investigate what kind of ligand(s) binds to the START domain of PDF2, Arabidopsis cell lines were generated by overexpressing either full-length PDF2 or mutants lacking the START domain. The recombinant proteins were tagged with a tandem affinity purification (TAP)-tag and TAP-tagged proteins were immunoprecipitated from cell lysates. After using mass-spectrometry (MS)-based proteomics to confirm the purification of PDF2, a liquid-chromatography MS (LC-MS) platform that targeted more than 240 lipid species was used to identify lipids co-purifying with PDF2. This analysis revealed that 12 lipid species were at least four-fold enriched in samples containing PDF2 compared to the mutant protein lacking the START domain. The lipid species with the highest degree of enrichment (seven to 12-fold) was lyso-PC.
To confirm that lyso-PC binds to the START domain, a fusion between the START domain and maltose binding protein (MBP) was expressed in Escherichia coli. As a control, a similar fusion protein mutated in the presumed lipid-binding pocket of the START domain was also expressed. After MBP cleavage, the interaction between the START domains and lyso-PC was studied by analyzing changes in fluorescence in microscale thermophoresis (MST) assays. In this assay, both the wild-type (WT) START domain and the mutant form bound phosphatidylcholine (PC) liposomes with the same affinity. However, in a reconstituted system where liposomes were doped with lyso-PC, the WT START domain bound the liposomes with an affinity c. 12-fold higher than that of the mutant.
In summary, purification of PDF2 from Arabidopsis cell cultures revealed enrichment in lyso-PCs, and direct interaction between the PDF2 START domain and lyso-PCs was verified by in vitro binding assays using liposomes. This suggested to the authors that PDF2 functions as a lyso-PC sensor via its START domain.
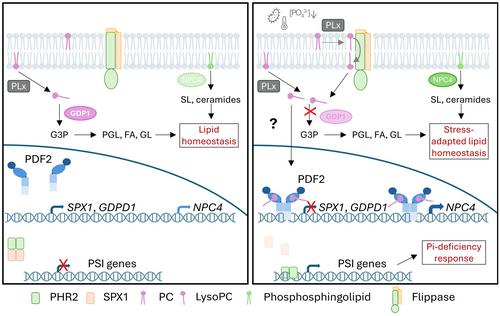
DNA-PDF2 electrophoretic mobility shift assays (EMSA) further demonstrated that PDF2 binds an octameric DNA promoter motif implicated in phospholipid remodeling and the phosphate starvation response (Wojciechowska et al.). Perfect matches were found in the promoters or 5′-UTR regions of glycerophosphodiester phosphodiesterase (GDPD1), the SPX domain-containing nuclear protein SPX1, and a nonspecific phospholipase C (NPC4), amongst other genes. Subsequent quantitative reverse transcription polymerase chain reaction analysis demonstrated that PDF2 is a transcriptional repressor for GDPD1 and SPX1. While a clear repression or activation pattern was not found for NPC4 in response to PDF2, this gene is highly upregulated during phosphate starvation and PDF2 might contribute to the modulation of this response (Yang et al., 2021; Wojciechowska et al.). Together with the results of lipid-binding assays, this suggests that Arabidopsis PDF2 can control gene expression by sensing lipids in an analogous way as nuclear receptors operate in animals.
Root length was found to be reduced in pdf2 mutants and enlarged in PDF2 overexpressors, which suggested that PDF2 is important for elongation growth. As a possible mechanism, it is suggested that PDF2 transcriptionally regulates the synthesis of phospholipids required for membrane elongation. It is well known that phosphate starvation triggers a response where primary root growth is arrested and compensated by lateral root formation and root hair growth (Péret et al., 2011). Thus, by sensing the phospholipid status in the epidermis when inorganic phosphate is limited, the activity of PDF2 may adjust the synthesis of membrane lipids to trigger a root growth response (Wojciechowska et al., 2024).
What could be the physiological implications of lyso-PCs binding to PDF2 (Fig. 1)? Under phosphate starvation, different phospholipases are activated to trigger the replacement of phospholipids by galactolipids and/or the accumulation of triacylglycerols (TAGs), which allows the plant cells to recycle phosphate (Roychowdhury et al., 2023). This ultimately results in a marked decrease in lyso-PC levels, but transient increases in lysolipid levels have been documented for other plant responses requiring membrane remodeling (Roychowdhury et al., 2023). In addition, extracellular lyso-PC is actively taken up into the roots of Arabidopsis by the lipid flippase ALA10, which might allow the plant to sense the availability of inorganic phosphate from decaying humus in the soil (Poulsen et al., 2015). It has long been known that lyso-PCs activate the plasma membrane H+-ATPase (Palmgren & Sommarin, 1989), which provides the driving force for uptake of phosphate in root epidermal cells, and induction of phosphate transporters in response to lyso-PC levels has been documented in arbuscular mycorrhizal symbioses in tomato and during Zn starvation responses in Arabidopsis (Drissner et al., 2007; Kisko et al., 2018). Thus, the results from Wojciechowska et al. suggest that fast changes in lyso-PC levels due to PC degradation and flippase internalization may activate not only the plasma membrane H+-ATPase, but also the phosphate starvation response via PDF2. In this way, PDF2 provides a link between the metabolic states and cell growth and differentiation.
An open question in the work of Wojciechowska et al. is how lyso-PC and PDF2 meet in the cell. While no answer to this question is provided, Wojciechowska et al. suggest that a co-repressor acts with PDF2. One possibility is that the co-repressor binds lyso-PC before travelling to the nucleus and delivers the lipid to PDF2. Alternatively, cytosolic PDF2 would directly bind lyso-PC and then travel to the nucleus to exert its function.
Last but not least, lyso-PC is proposed to be a signaling lipid, not only during phosphate starvation, but also in other environmental stresses (cold, heat, pathogens) (Hou et al., 2016). It has been suggested that the phosphate starvation response overlaps with the responses to these other stresses by use of shared transcription factors (Baek et al., 2017). Thus, the work of Wojciechowska et al. generates several questions. What other plant HD-Zip IV transcription factors sense lyso-PC? Is PDF2 involved in lyso-PC-triggered responses to other stresses? Several lipid species different from lyso-PC were identified in lipidomics analysis of full-length PDF2 purified from plant cells. Can the START domain of PDF2 bind different lipids with different affinities? Is this used as a mechanism to sense different types of stresses that might result in distinctly altered lipid ratios?
期刊介绍:
New Phytologist is an international electronic journal published 24 times a year. It is owned by the New Phytologist Foundation, a non-profit-making charitable organization dedicated to promoting plant science. The journal publishes excellent, novel, rigorous, and timely research and scholarship in plant science and its applications. The articles cover topics in five sections: Physiology & Development, Environment, Interaction, Evolution, and Transformative Plant Biotechnology. These sections encompass intracellular processes, global environmental change, and encourage cross-disciplinary approaches. The journal recognizes the use of techniques from molecular and cell biology, functional genomics, modeling, and system-based approaches in plant science. Abstracting and Indexing Information for New Phytologist includes Academic Search, AgBiotech News & Information, Agroforestry Abstracts, Biochemistry & Biophysics Citation Index, Botanical Pesticides, CAB Abstracts®, Environment Index, Global Health, and Plant Breeding Abstracts, and others.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


