{"title":"Correction to ‘Life history mediates the trade-offs among different components of demographic resilience’","authors":"","doi":"10.1111/ele.14468","DOIUrl":null,"url":null,"abstract":"<p>Capdevila P, Stott I, Cant J, Beger M, Rowlands G, Grace M, Salguero-Gómez R. (2022) Life history mediates the trade-offs among different components of demographic resilience. <i>Ecology Letters</i>, 25(6), 1566–1579. https://doi.org/10.1111/ele.14004</p><p>The authors note a mistake in the calculation of resistance in the methods section, which alters the interpretation of some of the results. In the manuscript, we applied a correction to calculation of resistance (<span></span><math>\n <semantics>\n <mrow>\n <msub>\n <munder>\n <mi>ρ</mi>\n <mo>_</mo>\n </munder>\n <mn>1</mn>\n </msub>\n </mrow>\n <annotation>$$ {\\underset{\\_}{\\rho}}_1 $$</annotation>\n </semantics></math>) in Equation (3) by subtracting the first step attenuation from 1 (1 − <span></span><math>\n <semantics>\n <mrow>\n <msub>\n <munder>\n <mi>ρ</mi>\n <mo>_</mo>\n </munder>\n <mn>1</mn>\n </msub>\n </mrow>\n <annotation>$$ {\\underset{\\_}{\\rho}}_1 $$</annotation>\n </semantics></math>). Such subtraction in the formula was an error, and the correct calculation should simply be <span></span><math>\n <semantics>\n <mrow>\n <msub>\n <munder>\n <mi>ρ</mi>\n <mo>_</mo>\n </munder>\n <mn>1</mn>\n </msub>\n </mrow>\n <annotation>$$ {\\underset{\\_}{\\rho}}_1 $$</annotation>\n </semantics></math> where values close to 1 correspond to high resistance and 0 to low resistance.</p><p>In Figure 2 of the original manuscript, we showed that the phylogenetic signal for resistance was 0.48 ± 0.26 (mean ± SE) in animals and 0.02 ± 0.04 in plants. When applying the correct calculation of resistance the phylogenetic signal remains virtually unaltered, with values of 0.45 ± 0.25 and 0.03 ± 0.04, respectively.</p><p>In Figure 3 of the original manuscript, we showed that resistance and recovery time were positively associated in animals and slightly, negatively associated in plants. Also, resistance and compensation were positively associated in animals and plants. When applying the correct calculation of resistance, the same associations hold, but the correlation values are inverted. That is, resistance and recovery time are negatively correlated in animals (Figure 3a) and positively correlated in plants (Figure 3d), while resistance and compensation are negatively correlated for both animals (Figure 3d) and plants (Figure 3e).</p><p>In Figure 4 of the original version of our paper, we showed that resistance was negatively associated with generation time in animals, while it was positively associated in plants. We also showed that resistance was positively associated with reproductive output for both plants and animals. When applying the correct calculation of resistance, we again show that the associations hold, but naturally they are reversed (Figure 4). Resistance is positively associated with generation time in animals and negatively associated in plants (Figure 4b), and resistance is negatively associated with reproductive output in both plants and animals (Figure 4e).</p><p>In the original supplementary analyses, we showed that most of the correlations among the life history traits and the resilience components were not spurious. When applying the correct calculation of resistance, there are still discrepancies between the simulated and the natural populations in their association of resistance with life history traits. The only difference is that the relationships with resistance are inverted (Figure S1).</p><p>In the original supplementary analyses, we showed resistance was independent of body dimension for both animals and plants, with the slopes of these correlations showing no clear trend. This pattern holds for the updated version of the plot but with the resistance axis reversed (Figure S2b,e).</p><p>In Figure S3 of the original supplementary materials, we showed that resistance did not have any clear association with the Raunkiær life forms. The updated version shows the same patterns, but with the resistance axes reversed (Figure S3, middle panel).</p><p>In Figure S4 of the original supplementary materials, we showed that there was a high variation in the compensation, resistance and recovery time among the different conservation statuses of the species (Figure S4). These results remain unaltered, but the resistance axes are reversed (Figure S4, middle panels).</p>","PeriodicalId":161,"journal":{"name":"Ecology Letters","volume":"27 7","pages":""},"PeriodicalIF":7.6000,"publicationDate":"2024-07-22","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1111/ele.14468","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"Ecology Letters","FirstCategoryId":"93","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1111/ele.14468","RegionNum":1,"RegionCategory":"环境科学与生态学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"ECOLOGY","Score":null,"Total":0}
引用次数: 0
Abstract
Capdevila P, Stott I, Cant J, Beger M, Rowlands G, Grace M, Salguero-Gómez R. (2022) Life history mediates the trade-offs among different components of demographic resilience. Ecology Letters, 25(6), 1566–1579. https://doi.org/10.1111/ele.14004
The authors note a mistake in the calculation of resistance in the methods section, which alters the interpretation of some of the results. In the manuscript, we applied a correction to calculation of resistance () in Equation (3) by subtracting the first step attenuation from 1 (1 − ). Such subtraction in the formula was an error, and the correct calculation should simply be where values close to 1 correspond to high resistance and 0 to low resistance.
In Figure 2 of the original manuscript, we showed that the phylogenetic signal for resistance was 0.48 ± 0.26 (mean ± SE) in animals and 0.02 ± 0.04 in plants. When applying the correct calculation of resistance the phylogenetic signal remains virtually unaltered, with values of 0.45 ± 0.25 and 0.03 ± 0.04, respectively.
In Figure 3 of the original manuscript, we showed that resistance and recovery time were positively associated in animals and slightly, negatively associated in plants. Also, resistance and compensation were positively associated in animals and plants. When applying the correct calculation of resistance, the same associations hold, but the correlation values are inverted. That is, resistance and recovery time are negatively correlated in animals (Figure 3a) and positively correlated in plants (Figure 3d), while resistance and compensation are negatively correlated for both animals (Figure 3d) and plants (Figure 3e).
In Figure 4 of the original version of our paper, we showed that resistance was negatively associated with generation time in animals, while it was positively associated in plants. We also showed that resistance was positively associated with reproductive output for both plants and animals. When applying the correct calculation of resistance, we again show that the associations hold, but naturally they are reversed (Figure 4). Resistance is positively associated with generation time in animals and negatively associated in plants (Figure 4b), and resistance is negatively associated with reproductive output in both plants and animals (Figure 4e).
In the original supplementary analyses, we showed that most of the correlations among the life history traits and the resilience components were not spurious. When applying the correct calculation of resistance, there are still discrepancies between the simulated and the natural populations in their association of resistance with life history traits. The only difference is that the relationships with resistance are inverted (Figure S1).
In the original supplementary analyses, we showed resistance was independent of body dimension for both animals and plants, with the slopes of these correlations showing no clear trend. This pattern holds for the updated version of the plot but with the resistance axis reversed (Figure S2b,e).
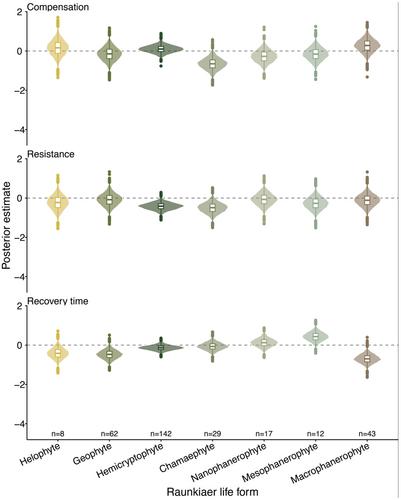
In Figure S3 of the original supplementary materials, we showed that resistance did not have any clear association with the Raunkiær life forms. The updated version shows the same patterns, but with the resistance axes reversed (Figure S3, middle panel).
In Figure S4 of the original supplementary materials, we showed that there was a high variation in the compensation, resistance and recovery time among the different conservation statuses of the species (Figure S4). These results remain unaltered, but the resistance axes are reversed (Figure S4, middle panels).
期刊介绍:
Ecology Letters serves as a platform for the rapid publication of innovative research in ecology. It considers manuscripts across all taxa, biomes, and geographic regions, prioritizing papers that investigate clearly stated hypotheses. The journal publishes concise papers of high originality and general interest, contributing to new developments in ecology. Purely descriptive papers and those that only confirm or extend previous results are discouraged.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


