Exploring solute binding proteins in Pseudomonas aeruginosa that bind to γ-aminobutyrate and 5-aminovalerate and their role in activating sensor kinases
{"title":"Exploring solute binding proteins in Pseudomonas aeruginosa that bind to γ-aminobutyrate and 5-aminovalerate and their role in activating sensor kinases","authors":"Jean Paul Cerna-Vargas, Tino Krell","doi":"10.1002/mbo3.1415","DOIUrl":null,"url":null,"abstract":"<p>The standard method of receptor activation involves the binding of signals or signal-loaded solute binding proteins (SBPs) to sensor domains. Many sensor histidine kinases (SHKs), which are activated by SBP binding, are encoded adjacent to their corresponding sbp gene. We examined three SBPs of <i>Pseudomonas aeruginosa</i> PAO1, encoded near the genes for the AgtS (PA0600) and AruS (PA4982) SHKs, to determine how common this arrangement is. Ligand screening and microcalorimetric studies revealed that the SBPs PA0602 and PA4985 preferentially bind to GABA (KD = 2.3 and 0.58 μM, respectively), followed by 5-aminovalerate (KD = 30 and 1.6 μM, respectively) and ethanoldiamine (KD = 2.3 and 0.58 μM, respectively). In contrast, AgtB (PA0604) exclusively recognizes 5-aminovaleric acid (KD = 2.9 μM). However, microcalorimetric titrations did not show any binding between the AgtS sensor domain and AgtB or PA0602, regardless of the presence of ligands. Similarly, bacterial two-hybrid assays did not demonstrate an interaction between PA4985 and the AruS sensor domain. Therefore, sbp and shk genes located nearby are not always functionally linked. We previously identified PA0222 as a GABA-specific SBP. The presence of three SBPs for GABA may be linked to GABA's role as a trigger for <i>P. aeruginosa</i> virulence.</p>","PeriodicalId":18573,"journal":{"name":"MicrobiologyOpen","volume":"13 3","pages":""},"PeriodicalIF":4.6000,"publicationDate":"2024-05-23","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1002/mbo3.1415","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"MicrobiologyOpen","FirstCategoryId":"99","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1002/mbo3.1415","RegionNum":3,"RegionCategory":"生物学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q2","JCRName":"MICROBIOLOGY","Score":null,"Total":0}
引用次数: 0
Abstract
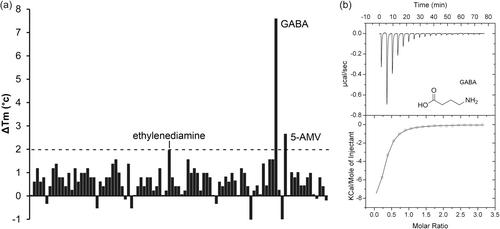
The standard method of receptor activation involves the binding of signals or signal-loaded solute binding proteins (SBPs) to sensor domains. Many sensor histidine kinases (SHKs), which are activated by SBP binding, are encoded adjacent to their corresponding sbp gene. We examined three SBPs of Pseudomonas aeruginosa PAO1, encoded near the genes for the AgtS (PA0600) and AruS (PA4982) SHKs, to determine how common this arrangement is. Ligand screening and microcalorimetric studies revealed that the SBPs PA0602 and PA4985 preferentially bind to GABA (KD = 2.3 and 0.58 μM, respectively), followed by 5-aminovalerate (KD = 30 and 1.6 μM, respectively) and ethanoldiamine (KD = 2.3 and 0.58 μM, respectively). In contrast, AgtB (PA0604) exclusively recognizes 5-aminovaleric acid (KD = 2.9 μM). However, microcalorimetric titrations did not show any binding between the AgtS sensor domain and AgtB or PA0602, regardless of the presence of ligands. Similarly, bacterial two-hybrid assays did not demonstrate an interaction between PA4985 and the AruS sensor domain. Therefore, sbp and shk genes located nearby are not always functionally linked. We previously identified PA0222 as a GABA-specific SBP. The presence of three SBPs for GABA may be linked to GABA's role as a trigger for P. aeruginosa virulence.
期刊介绍:
MicrobiologyOpen is a peer reviewed, fully open access, broad-scope, and interdisciplinary journal delivering rapid decisions and fast publication of microbial science, a field which is undergoing a profound and exciting evolution in this post-genomic era.
The journal aims to serve the research community by providing a vehicle for authors wishing to publish quality research in both fundamental and applied microbiology. Our goal is to publish articles that stimulate discussion and debate, as well as add to our knowledge base and further the understanding of microbial interactions and microbial processes.
MicrobiologyOpen gives prompt and equal consideration to articles reporting theoretical, experimental, applied, and descriptive work in all aspects of bacteriology, virology, mycology and protistology, including, but not limited to:
- agriculture
- antimicrobial resistance
- astrobiology
- biochemistry
- biotechnology
- cell and molecular biology
- clinical microbiology
- computational, systems, and synthetic microbiology
- environmental science
- evolutionary biology, ecology, and systematics
- food science and technology
- genetics and genomics
- geobiology and earth science
- host-microbe interactions
- infectious diseases
- natural products discovery
- pharmaceutical and medicinal chemistry
- physiology
- plant pathology
- veterinary microbiology
We will consider submissions across unicellular and cell-cluster organisms: prokaryotes (bacteria, archaea) and eukaryotes (fungi, protists, microalgae, lichens), as well as viruses and prions infecting or interacting with microorganisms, plants and animals, including genetic, biochemical, biophysical, bioinformatic and structural analyses.
The journal features Original Articles (including full Research articles, Method articles, and Short Communications), Commentaries, Reviews, and Editorials. Original papers must report well-conducted research with conclusions supported by the data presented in the article. We also support confirmatory research and aim to work with authors to meet reviewer expectations.
MicrobiologyOpen publishes articles submitted directly to the journal and those referred from other Wiley journals.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


