Changes in species richness with climate change in subalpine communities are dependent on regional environmental conditions and local functional composition
Richard Michalet, Ida Delpy, Auriane Eisenberg, Colin Hostein, Blaise Touzard, Nicolas Gross, Julien Pottier
{"title":"Changes in species richness with climate change in subalpine communities are dependent on regional environmental conditions and local functional composition","authors":"Richard Michalet, Ida Delpy, Auriane Eisenberg, Colin Hostein, Blaise Touzard, Nicolas Gross, Julien Pottier","doi":"10.1111/jvs.13254","DOIUrl":null,"url":null,"abstract":"<p>Climate change is considered to severely threaten overall earth biodiversity, both nowadays and in the near future. However, the evidence for sharp decreases in diversity has mainly been found in marine and freshwater systems and for animals (Bellwood et al., <span>2004</span>; Wilson et al., <span>2005</span>; Reid et al., <span>2019</span>; Albert et al., <span>2021</span>), but less so in terrestrial systems and for plant species which show a higher resistance to climate change than originally predicted (Thuiller et al., <span>2005</span>; Bertrand et al., <span>2011</span>; Cheaib et al., <span>2012</span>; Lenoir, <span>2022</span>).</p><p>Alpine plant communities are considered the most sensitive terrestrial system for biodiversity changes because of the steepness of ecological gradients and the isolation of most alpine summits (Körner, <span>2003</span>; Anthelme & Lavergne, <span>2018</span>). However, changes in species richness are highly dependent on the environmental context in terms of elevation, regional climates, soil types, snow cover duration and human pressure (Kammer et al., <span>2007</span>; Scherrer & Körner, <span>2011</span>; Rumpf et al., <span>2018</span>; Nicklas et al., <span>2021</span>). Pioneer resurvey studies have shown that species richness has increased at the high alpine and nival belts, probably due to climate warming (Grabherr et al., <span>1994</span>; Gottfried et al., <span>2012</span>; Pauli et al., <span>2012</span>; Steinbauer et al., <span>2018</span>), but the few studies conducted at the subalpine belt have rather observed decreases in species richness probably due to increasing competition with climate warming (Vittoz et al., <span>2009</span>; Alexander et al., <span>2015</span>; Nicklas et al., <span>2021</span>; Michalet et al., <span>2024</span>). Different regional climates may also trigger different community responses because competition is more likely to increase with warming in wet climates, whereas facilitation is expected to increase with increasing drought in drier climates, with opposite consequences for species richness (Choler et al., <span>2001</span>; Callaway et al., <span>2002</span>; Michalet et al., <span>2014</span>). For example, comparing changes in alpine vegetation between the temperate climate of the Central Alps, the submediterranean climate of the Apennines and the drier Mediterranean climate of Crete, Steinbauer et al. (<span>2022</span>) found a thermophilization of the vegetation in the two northern wet mountains only, with a decline of cold-adapted cushion plants in the Alps. In addition, different mountain regions can further show dissimilar species pools, which is well recognized as a source of contingencies in community responses (Lessard et al., <span>2012</span>).</p><p>Changes in species richness are also highly dependent on snow cover duration varying across space and time, that is, along mesotopography gradients and with climate change, respectively (Matteodo et al., <span>2016</span>). Choler (<span>2015</span>, <span>2018</span>) highlighted that the positive effect of an extended growing season due to decreasing snow cover can be counteracted by the detrimental effects of increasing the number of freezing days, with contrasting consequences for species’ responses along mesotopographic gradients depending on species’ functional strategies (see also Wipf et al., <span>2009</span>). Sandvik and Odland (<span>2014</span>) assessed changes in Alpine snowbeds from Norway over a 30-year-long period and found an increase in species richness in both dry and wet snowbeds, mostly because of the invasion by shrubs, graminoids and herbs. However, drought-intolerant species were strongly reduced. Daniëls et al. (<span>2011</span>) found high changes in several snowbed communities from Greenland with an overall increase in species richness due to increasing drought. Carbognani et al. (<span>2014</span>) assessed vegetation changes in an alpine snowbed from the Italian Alps and found a high internal dynamic due to increased competition, but also an overall increase in species richness. In contrast, modeling the effects of changes in snow cover duration on arctic plant communities, Niittynen et al. (<span>2018</span>) found that a 20%–30% decrease in snow cover duration might fully compensate for the increase in species richness due to warming, potentially inducing important losses of vascular plants, lichens and bryophytes. Niittynen et al. (<span>2020</span>) also found with modeling that decreasing snow cover, together with warming temperatures, can substantially modify plant functional traits, with future plant communities projected to be occupied by taller plants with larger leaves and faster growth, and, thus, likely inducing an increase in competition and decrease in species richness.</p><p>Plant community richness is known to be highly dependent on the functional composition of plant communities closely linked to biotic interactions, i.e., facilitation and competition (Grime, <span>1973</span>; Michalet et al., <span>2006</span>; Michalet et al., <span>2023</span>; Munoz et al., <span>2023</span>). Because functional composition also varies with environmental conditions, in particular climate and soil types (Díaz et al., <span>2016</span>; Joswig et al., <span>2022</span>), the effect of global change on diversity is likely to be mediated by changes in functional composition (Bjorkman et al., <span>2018</span>; Niittynen et al., <span>2020</span>; Bektas et al., <span>2023</span>). Although it is not easy to separate the effects of local environmental conditions from those of functional composition, the latter can also influence changes in diversity with climate change due to differences in the relative frequency and abundance of functional strategies across mountain systems. For example, a high relative frequency of species intolerant to competition (i.e., short exploitative or short conservative species sensu Michalet et al., <span>2023</span>) may enhance variation in species richness through changes in competitive responses with climate change. Alternatively, a high relative abundance of competitors (i.e., tall exploitative or tall conservative species sensu Michalet et al., <span>2023</span>) may enhance variation in competitive effects of community dominants. Thus, since alpine systems with harsh physical conditions (e.g., continental and/or dry climates) are more likely to be dominated by short species intolerant to competition (Joswig et al., <span>2022</span>; Michalet et al., <span>2023</span>), we may expect strong decreases in diversity as environmental conditions improve with climate change in such mountain range due to the high local potential for competitive exclusion (i.e., due to the abundance of species intolerant to competition that could be excluded). Alternatively, in milder environmental conditions, the increase in competitive exclusion should be less pronounced due to the lack of species likely to be excluded. As a second alternative, in the case of pejoration of environmental conditions with climate change, increases in diversity are more likely to be the strongest in mild environmental conditions due to decreasing competitive effects there and even loss of species in the less favorable mountain range already subjected to harsh environmental conditions.</p><p>In this study we aim to compare changes in species richness with climate change in two contrasting mountainous areas, a dry continental mountain range from the southwestern Alps (Alpe d'Huez, French Alps) and a wet oceanic mountain range from the Massif Central (France). The higher physical stress of the former mountain range is strengthened by the occurrence of calcareous rocks (in addition to siliceous rocks), whereas the more favorable physical conditions of the latter are even improved by a homogenous volcanic substrate characterized by deeper soils with higher water availability than calcareous rocks (Michalet et al., <span>2002</span>; Nemer et al., <span>2021</span>). The two systems are very likely to also show contrasting functional compositions, due to the occurrence of dry short calcareous grasslands and tall siliceous heaths in the former and wet tall forb communities in the latter (Michalet & Philippe, <span>1993</span>). Vegetation composition was assessed in 1997 and 2017–2018 in the Alps and 1988–1989 and 2022 in the Massif Central and functional composition was assessed through contemporary trait measurements in the two mountain ranges. We used multivariate analyses for quantifying correlations between changes in species richness, functional composition and environmental conditions in the two mountain ranges separately. We also quantified changes in climatic conditions and species richness between the ancient and recent vegetation surveys in each mountain range. We addressed four main questions: (i) did climate change similarly in the two mountain ranges, in terms of temperatures, water balance, snow cover duration and irradiance? (ii) What are the differences in functional composition between the two mountain ranges? (iii) What are the drivers of variation in species richness along spatial environmental gradients in the two mountain ranges? (iv) Did species richness show contrasting variations with climate change in the two mountain ranges?</p><p>At both sites during the 1990–2020 period, summer hydric balance significantly decreased (<i>r</i> = −0.39 and −0.55, <i>p</i> = 0.03 and 0.001, for the Alpe d'Huez and Sancy sites, respectively, Figure 2a,b) and summer mean of maximum temperatures significantly increased (<i>r</i> = 0.56 and 0.43, <i>p</i> = 0.001 and 0.015, for the Alpe d'Huez and Sancy sites, respectively, Figure 2c,d). Summer sum of global irradiance only significantly increased at the Sancy site (<i>r</i> = 0.39, <i>p</i> = 0.03, Figure 2f), whereas summer mean nebulosity did not significantly vary at the Alpe d'Huez site (<i>p</i> = 0.38, Figure 2e). In contrast, snow cover duration significantly decreased at the Alpe d'Huez site (<i>r</i> = −0.53, <i>p</i> = 0.002, Figure 2g), but did not significantly vary at the Sancy site (<i>p</i> = 0.45, Figure 2h).</p><p>The PCA conducted on the traits of the species of both sites showed two main axes of functional variation: the first axis correlated to the size traits (height, lateral spread and leaf area) and the second axis correlated to growth traits (SLA and LDMC). Thus, four main functional syndromes may be observed, tall conservative species at the upper right of the diagram (e.g., <i>Rhododendron ferrugineum</i>, only present at the Alpe d'Huez site, mostly on wet oligotrophic siliceous soils), tall exploitative species at the lower right of the diagram (e.g., <i>Heracleum sphondylium</i>, only present at the Sancy site, mostly in tall megaphorbias on wet eutrophic soils), short conservative species at the upper left of the diagram (e.g., <i>Globularia cordifolia</i>, only present at the Alpe d'Huez site, mostly on dry calcareous ridges) and short exploitative species at the lower left of the diagram (e.g., <i>Potentilla aurea</i>, present at both sites, mostly in short wet grasslands from snowbeds) (Figure 3). Species from both sites had very different overall positions in the diagram, with primarily much taller species at the Sancy than Alpe d'Huez site (highly significant site effect on the PCA 1 axis; <i>F</i> = 43.1, <i>p</i> < 0.001). Species from the Sancy site were also more exploitative than species from the Alpe d'Huez site (highly significant site effect on the PCA 2 axis; <i>F</i> = 19.1, <i>p</i> < 0.001).</p><p>At the Alpe d'Huez, there was, for both years of sampling, a highly significant unimodal relationship between species richness and the position of the plots on the first axis of the CA conducted on vegetation composition (Figure 4 and see Appendix S2 showing that plot positions on the first axis of the CCA [and CA, as both diagrams were highly similar] were primarily correlated to water availability and vegetation cover). This means that, for both years of sampling, species richness was the highest at intermediate position along the first CA axis in plant communities of intermediate cover and biomass, i.e., mesic calcareous grasslands dominated by <i>Carex sempervirens</i>, but lower in short dry calcareous grasslands dominated by <i>Globularia cordifolia</i> and the lowest in the wettest communities on siliceous rocks with the highest cover and biomass (heathlands dominated by ericaceous shrubs). There was no statistically significant difference in plot scores on the first CA axis between the two years of sampling (<i>F</i><sub>1,188</sub> = 1.06, <i>p</i> = 0.305; Figure 4). There was a statistically significant difference in species richness between the two years of sampling (<i>F</i><sub>1,188</sub> = 99.18, <i>p</i> < 0.001), with on average a 10% decrease from 1997 to 2017–2018. Finally, we found a statistically significant interaction between time and clusters (<i>F</i><sub>11,177</sub> = 2.46, <i>p</i> = 0.008; Figure 5a). Considering Bonferroni-adjusted <i>p</i>-values, the changes in species richness between the two years of sampling were significant only for calcareous short grasslands, calcareous <i>Juniper</i> heathlands, <i>Vaccinium</i> heathlands from hollows, and siliceous short grasslands.</p><p>At the Sancy site, there was, in both years of sampling, a highly significant linear decrease in species richness along the second axis of the CA conducted on vegetation composition (Figure 6 and see Appendixes S3 and S4B showing that plot position on the second axis of the CA was primarily correlated to the radiation index, i.e., global radiation, and, thus, exposure). This means that, in both years of sampling, the highest species richness was found in communities with a southern exposure where irradiance was the highest, that is, tall mesotrophic meadows dominated by <i>Patzkea paniculata</i> or <i>Calamagrostis arundinacea</i>, and the lowest species richness was found in communities with a northern exposure where irradiance was the lowest, i.e., tall eutrophic megaphorbias and short grasslands from snowbeds dominated by <i>Nardus stricta</i>. There was a statistically significant difference in plot scores along the CA axis 2 between the two years of sampling (<i>F</i>1,156 = 12.35, <i>p</i> < 0.001; Figure 6). This means that, with climate change, the relative importance of species with a southern exposure increased, whereas the importance of species with a northern exposure decreased. There was a statistically significant difference in species richness between the two years of sampling (<i>F</i><sub>1,156</sub> = 21.464, <i>p</i> < 0.001, Figure 6), with an overall increase of 10% from 1988–1989 to 2022. Finally, we found no statistically significant interaction between time and clusters (<i>F</i><sub>12,144</sub> = 1.74, <i>p</i> = 0.065, Figure 5b). However, the analysis confirms a clear cluster effect on species richness <i>F</i><sub>12,144</sub> = 15.78, <i>p</i> < 0.001 and of the year of sampling <i>F</i><sub>1,144</sub> = 16.01, <i>p</i> < 0.001. In contrast, there was no relationship between species richness and the first CA axis (data not shown), that was significantly explained by the erosivity index (i.e., LS, see section 2. Materials and Methods) controlling soil fertility and nutrient availability (Appendix S4A).</p><p>In response to our first question, climate change induced a similar increase in summer temperature and decrease in water balance in the two subalpine systems, but snow cover duration decreased only in the dry semi-continental Alpe d'Huez site, whereas summer global radiations increased in the wet oceanic Sancy site only. In response to our second question, there were strong differences in functional composition between the two sites with taller and more exploitative species in the Sancy site than in the Alpe d'Huez site. In response to our third question, spatial variation in species richness was driven by different ecological processes at the two sites. In the Alps, species richness showed a unimodal relationship along a physical stress/community biomass gradient, whereas, in the Massif Central, species richness increased with increasing global radiation from north to south exposure. Finally, in response to our fourth question, there were significant changes in species richness with climate change at both sites, but in opposite directions, with a negative effect of climate change in the Alps but a positive effect in the Massif Central. These results suggest that variation in species richness with climate change in these two subalpine systems was highly dependent on the regional context.</p><p>Our results suggest that changes in species richness with recent climate change of subalpine communities from two mountain ranges having contrasting climatic (and bedrock) conditions depend on the interplay between the level of physical stress in each mountain range, their functional composition and different drivers of variation in species richness along spatial ecological gradients between the two mountain ranges, together with the specific climatic variations induced by climate change in each range. In the dry and semi-continental site of Alpe d'Huez dominated by short conservative species, climate change induced a decrease in species richness likely because spatial variation in species richness was primarily driven by variation in community biomass and because climate change occurred mostly at this site through a strong decrease in snow cover duration that increased community biomass and, thus, likely the competitive exclusion of short species. In contrast, in the wet oceanic Sancy site dominated by tall exploitative species, climate change induced an increase in species richness because variation in species richness is primarily driven by variation in global radiation that significantly increased with climate change, thus, inducing a positive light and warming effect for short species, in particular short exploitative species.</p><p>Richard Michalet did the sampling in 1997 and Richard Michalet and Blaise Touzard the sampling in 2017–2018 at the Alpe d'Huez site. Richard Michalet did the sampling in 1988–1989 and Richard Michalet, Blaise Touzard, Ida Delpy and Colin Hostein sampled vegetation composition in 2022. Auriane Eisenberg, Julien Pottier and Nicolas Gross sampled functional traits in 2022 at the Sancy site. Richard Michalet, Julien Pottier and Nicolas Gross ran the statistical analyses. Richard Michalet wrote the first draft of the manuscript and all authors contributed to editing it.</p><p>The authors have no conflicts of interest.</p>","PeriodicalId":49965,"journal":{"name":"Journal of Vegetation Science","volume":"35 2","pages":""},"PeriodicalIF":2.2000,"publicationDate":"2024-04-16","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1111/jvs.13254","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"Journal of Vegetation Science","FirstCategoryId":"93","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1111/jvs.13254","RegionNum":3,"RegionCategory":"环境科学与生态学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q2","JCRName":"ECOLOGY","Score":null,"Total":0}
引用次数: 0
Abstract
Climate change is considered to severely threaten overall earth biodiversity, both nowadays and in the near future. However, the evidence for sharp decreases in diversity has mainly been found in marine and freshwater systems and for animals (Bellwood et al., 2004; Wilson et al., 2005; Reid et al., 2019; Albert et al., 2021), but less so in terrestrial systems and for plant species which show a higher resistance to climate change than originally predicted (Thuiller et al., 2005; Bertrand et al., 2011; Cheaib et al., 2012; Lenoir, 2022).
Alpine plant communities are considered the most sensitive terrestrial system for biodiversity changes because of the steepness of ecological gradients and the isolation of most alpine summits (Körner, 2003; Anthelme & Lavergne, 2018). However, changes in species richness are highly dependent on the environmental context in terms of elevation, regional climates, soil types, snow cover duration and human pressure (Kammer et al., 2007; Scherrer & Körner, 2011; Rumpf et al., 2018; Nicklas et al., 2021). Pioneer resurvey studies have shown that species richness has increased at the high alpine and nival belts, probably due to climate warming (Grabherr et al., 1994; Gottfried et al., 2012; Pauli et al., 2012; Steinbauer et al., 2018), but the few studies conducted at the subalpine belt have rather observed decreases in species richness probably due to increasing competition with climate warming (Vittoz et al., 2009; Alexander et al., 2015; Nicklas et al., 2021; Michalet et al., 2024). Different regional climates may also trigger different community responses because competition is more likely to increase with warming in wet climates, whereas facilitation is expected to increase with increasing drought in drier climates, with opposite consequences for species richness (Choler et al., 2001; Callaway et al., 2002; Michalet et al., 2014). For example, comparing changes in alpine vegetation between the temperate climate of the Central Alps, the submediterranean climate of the Apennines and the drier Mediterranean climate of Crete, Steinbauer et al. (2022) found a thermophilization of the vegetation in the two northern wet mountains only, with a decline of cold-adapted cushion plants in the Alps. In addition, different mountain regions can further show dissimilar species pools, which is well recognized as a source of contingencies in community responses (Lessard et al., 2012).
Changes in species richness are also highly dependent on snow cover duration varying across space and time, that is, along mesotopography gradients and with climate change, respectively (Matteodo et al., 2016). Choler (2015, 2018) highlighted that the positive effect of an extended growing season due to decreasing snow cover can be counteracted by the detrimental effects of increasing the number of freezing days, with contrasting consequences for species’ responses along mesotopographic gradients depending on species’ functional strategies (see also Wipf et al., 2009). Sandvik and Odland (2014) assessed changes in Alpine snowbeds from Norway over a 30-year-long period and found an increase in species richness in both dry and wet snowbeds, mostly because of the invasion by shrubs, graminoids and herbs. However, drought-intolerant species were strongly reduced. Daniëls et al. (2011) found high changes in several snowbed communities from Greenland with an overall increase in species richness due to increasing drought. Carbognani et al. (2014) assessed vegetation changes in an alpine snowbed from the Italian Alps and found a high internal dynamic due to increased competition, but also an overall increase in species richness. In contrast, modeling the effects of changes in snow cover duration on arctic plant communities, Niittynen et al. (2018) found that a 20%–30% decrease in snow cover duration might fully compensate for the increase in species richness due to warming, potentially inducing important losses of vascular plants, lichens and bryophytes. Niittynen et al. (2020) also found with modeling that decreasing snow cover, together with warming temperatures, can substantially modify plant functional traits, with future plant communities projected to be occupied by taller plants with larger leaves and faster growth, and, thus, likely inducing an increase in competition and decrease in species richness.
Plant community richness is known to be highly dependent on the functional composition of plant communities closely linked to biotic interactions, i.e., facilitation and competition (Grime, 1973; Michalet et al., 2006; Michalet et al., 2023; Munoz et al., 2023). Because functional composition also varies with environmental conditions, in particular climate and soil types (Díaz et al., 2016; Joswig et al., 2022), the effect of global change on diversity is likely to be mediated by changes in functional composition (Bjorkman et al., 2018; Niittynen et al., 2020; Bektas et al., 2023). Although it is not easy to separate the effects of local environmental conditions from those of functional composition, the latter can also influence changes in diversity with climate change due to differences in the relative frequency and abundance of functional strategies across mountain systems. For example, a high relative frequency of species intolerant to competition (i.e., short exploitative or short conservative species sensu Michalet et al., 2023) may enhance variation in species richness through changes in competitive responses with climate change. Alternatively, a high relative abundance of competitors (i.e., tall exploitative or tall conservative species sensu Michalet et al., 2023) may enhance variation in competitive effects of community dominants. Thus, since alpine systems with harsh physical conditions (e.g., continental and/or dry climates) are more likely to be dominated by short species intolerant to competition (Joswig et al., 2022; Michalet et al., 2023), we may expect strong decreases in diversity as environmental conditions improve with climate change in such mountain range due to the high local potential for competitive exclusion (i.e., due to the abundance of species intolerant to competition that could be excluded). Alternatively, in milder environmental conditions, the increase in competitive exclusion should be less pronounced due to the lack of species likely to be excluded. As a second alternative, in the case of pejoration of environmental conditions with climate change, increases in diversity are more likely to be the strongest in mild environmental conditions due to decreasing competitive effects there and even loss of species in the less favorable mountain range already subjected to harsh environmental conditions.
In this study we aim to compare changes in species richness with climate change in two contrasting mountainous areas, a dry continental mountain range from the southwestern Alps (Alpe d'Huez, French Alps) and a wet oceanic mountain range from the Massif Central (France). The higher physical stress of the former mountain range is strengthened by the occurrence of calcareous rocks (in addition to siliceous rocks), whereas the more favorable physical conditions of the latter are even improved by a homogenous volcanic substrate characterized by deeper soils with higher water availability than calcareous rocks (Michalet et al., 2002; Nemer et al., 2021). The two systems are very likely to also show contrasting functional compositions, due to the occurrence of dry short calcareous grasslands and tall siliceous heaths in the former and wet tall forb communities in the latter (Michalet & Philippe, 1993). Vegetation composition was assessed in 1997 and 2017–2018 in the Alps and 1988–1989 and 2022 in the Massif Central and functional composition was assessed through contemporary trait measurements in the two mountain ranges. We used multivariate analyses for quantifying correlations between changes in species richness, functional composition and environmental conditions in the two mountain ranges separately. We also quantified changes in climatic conditions and species richness between the ancient and recent vegetation surveys in each mountain range. We addressed four main questions: (i) did climate change similarly in the two mountain ranges, in terms of temperatures, water balance, snow cover duration and irradiance? (ii) What are the differences in functional composition between the two mountain ranges? (iii) What are the drivers of variation in species richness along spatial environmental gradients in the two mountain ranges? (iv) Did species richness show contrasting variations with climate change in the two mountain ranges?
At both sites during the 1990–2020 period, summer hydric balance significantly decreased (r = −0.39 and −0.55, p = 0.03 and 0.001, for the Alpe d'Huez and Sancy sites, respectively, Figure 2a,b) and summer mean of maximum temperatures significantly increased (r = 0.56 and 0.43, p = 0.001 and 0.015, for the Alpe d'Huez and Sancy sites, respectively, Figure 2c,d). Summer sum of global irradiance only significantly increased at the Sancy site (r = 0.39, p = 0.03, Figure 2f), whereas summer mean nebulosity did not significantly vary at the Alpe d'Huez site (p = 0.38, Figure 2e). In contrast, snow cover duration significantly decreased at the Alpe d'Huez site (r = −0.53, p = 0.002, Figure 2g), but did not significantly vary at the Sancy site (p = 0.45, Figure 2h).
The PCA conducted on the traits of the species of both sites showed two main axes of functional variation: the first axis correlated to the size traits (height, lateral spread and leaf area) and the second axis correlated to growth traits (SLA and LDMC). Thus, four main functional syndromes may be observed, tall conservative species at the upper right of the diagram (e.g., Rhododendron ferrugineum, only present at the Alpe d'Huez site, mostly on wet oligotrophic siliceous soils), tall exploitative species at the lower right of the diagram (e.g., Heracleum sphondylium, only present at the Sancy site, mostly in tall megaphorbias on wet eutrophic soils), short conservative species at the upper left of the diagram (e.g., Globularia cordifolia, only present at the Alpe d'Huez site, mostly on dry calcareous ridges) and short exploitative species at the lower left of the diagram (e.g., Potentilla aurea, present at both sites, mostly in short wet grasslands from snowbeds) (Figure 3). Species from both sites had very different overall positions in the diagram, with primarily much taller species at the Sancy than Alpe d'Huez site (highly significant site effect on the PCA 1 axis; F = 43.1, p < 0.001). Species from the Sancy site were also more exploitative than species from the Alpe d'Huez site (highly significant site effect on the PCA 2 axis; F = 19.1, p < 0.001).
At the Alpe d'Huez, there was, for both years of sampling, a highly significant unimodal relationship between species richness and the position of the plots on the first axis of the CA conducted on vegetation composition (Figure 4 and see Appendix S2 showing that plot positions on the first axis of the CCA [and CA, as both diagrams were highly similar] were primarily correlated to water availability and vegetation cover). This means that, for both years of sampling, species richness was the highest at intermediate position along the first CA axis in plant communities of intermediate cover and biomass, i.e., mesic calcareous grasslands dominated by Carex sempervirens, but lower in short dry calcareous grasslands dominated by Globularia cordifolia and the lowest in the wettest communities on siliceous rocks with the highest cover and biomass (heathlands dominated by ericaceous shrubs). There was no statistically significant difference in plot scores on the first CA axis between the two years of sampling (F1,188 = 1.06, p = 0.305; Figure 4). There was a statistically significant difference in species richness between the two years of sampling (F1,188 = 99.18, p < 0.001), with on average a 10% decrease from 1997 to 2017–2018. Finally, we found a statistically significant interaction between time and clusters (F11,177 = 2.46, p = 0.008; Figure 5a). Considering Bonferroni-adjusted p-values, the changes in species richness between the two years of sampling were significant only for calcareous short grasslands, calcareous Juniper heathlands, Vaccinium heathlands from hollows, and siliceous short grasslands.
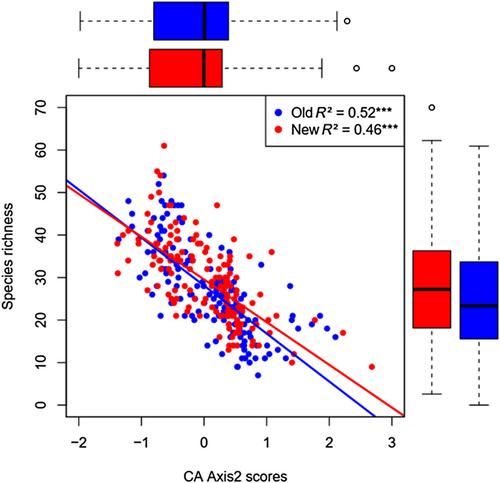
At the Sancy site, there was, in both years of sampling, a highly significant linear decrease in species richness along the second axis of the CA conducted on vegetation composition (Figure 6 and see Appendixes S3 and S4B showing that plot position on the second axis of the CA was primarily correlated to the radiation index, i.e., global radiation, and, thus, exposure). This means that, in both years of sampling, the highest species richness was found in communities with a southern exposure where irradiance was the highest, that is, tall mesotrophic meadows dominated by Patzkea paniculata or Calamagrostis arundinacea, and the lowest species richness was found in communities with a northern exposure where irradiance was the lowest, i.e., tall eutrophic megaphorbias and short grasslands from snowbeds dominated by Nardus stricta. There was a statistically significant difference in plot scores along the CA axis 2 between the two years of sampling (F1,156 = 12.35, p < 0.001; Figure 6). This means that, with climate change, the relative importance of species with a southern exposure increased, whereas the importance of species with a northern exposure decreased. There was a statistically significant difference in species richness between the two years of sampling (F1,156 = 21.464, p < 0.001, Figure 6), with an overall increase of 10% from 1988–1989 to 2022. Finally, we found no statistically significant interaction between time and clusters (F12,144 = 1.74, p = 0.065, Figure 5b). However, the analysis confirms a clear cluster effect on species richness F12,144 = 15.78, p < 0.001 and of the year of sampling F1,144 = 16.01, p < 0.001. In contrast, there was no relationship between species richness and the first CA axis (data not shown), that was significantly explained by the erosivity index (i.e., LS, see section 2. Materials and Methods) controlling soil fertility and nutrient availability (Appendix S4A).
In response to our first question, climate change induced a similar increase in summer temperature and decrease in water balance in the two subalpine systems, but snow cover duration decreased only in the dry semi-continental Alpe d'Huez site, whereas summer global radiations increased in the wet oceanic Sancy site only. In response to our second question, there were strong differences in functional composition between the two sites with taller and more exploitative species in the Sancy site than in the Alpe d'Huez site. In response to our third question, spatial variation in species richness was driven by different ecological processes at the two sites. In the Alps, species richness showed a unimodal relationship along a physical stress/community biomass gradient, whereas, in the Massif Central, species richness increased with increasing global radiation from north to south exposure. Finally, in response to our fourth question, there were significant changes in species richness with climate change at both sites, but in opposite directions, with a negative effect of climate change in the Alps but a positive effect in the Massif Central. These results suggest that variation in species richness with climate change in these two subalpine systems was highly dependent on the regional context.
Our results suggest that changes in species richness with recent climate change of subalpine communities from two mountain ranges having contrasting climatic (and bedrock) conditions depend on the interplay between the level of physical stress in each mountain range, their functional composition and different drivers of variation in species richness along spatial ecological gradients between the two mountain ranges, together with the specific climatic variations induced by climate change in each range. In the dry and semi-continental site of Alpe d'Huez dominated by short conservative species, climate change induced a decrease in species richness likely because spatial variation in species richness was primarily driven by variation in community biomass and because climate change occurred mostly at this site through a strong decrease in snow cover duration that increased community biomass and, thus, likely the competitive exclusion of short species. In contrast, in the wet oceanic Sancy site dominated by tall exploitative species, climate change induced an increase in species richness because variation in species richness is primarily driven by variation in global radiation that significantly increased with climate change, thus, inducing a positive light and warming effect for short species, in particular short exploitative species.
Richard Michalet did the sampling in 1997 and Richard Michalet and Blaise Touzard the sampling in 2017–2018 at the Alpe d'Huez site. Richard Michalet did the sampling in 1988–1989 and Richard Michalet, Blaise Touzard, Ida Delpy and Colin Hostein sampled vegetation composition in 2022. Auriane Eisenberg, Julien Pottier and Nicolas Gross sampled functional traits in 2022 at the Sancy site. Richard Michalet, Julien Pottier and Nicolas Gross ran the statistical analyses. Richard Michalet wrote the first draft of the manuscript and all authors contributed to editing it.
期刊介绍:
The Journal of Vegetation Science publishes papers on all aspects of plant community ecology, with particular emphasis on papers that develop new concepts or methods, test theory, identify general patterns, or that are otherwise likely to interest a broad international readership. Papers may focus on any aspect of vegetation science, e.g. community structure (including community assembly and plant functional types), biodiversity (including species richness and composition), spatial patterns (including plant geography and landscape ecology), temporal changes (including demography, community dynamics and palaeoecology) and processes (including ecophysiology), provided the focus is on increasing our understanding of plant communities. The Journal publishes papers on the ecology of a single species only if it plays a key role in structuring plant communities. Papers that apply ecological concepts, theories and methods to the vegetation management, conservation and restoration, and papers on vegetation survey should be directed to our associate journal, Applied Vegetation Science journal.
 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


