{"title":"Ontogenetic changes in ecophysiology are an understudied yet important component of plant adaptation","authors":"Erica H. Lawrence-Paul, Jesse R. Lasky","doi":"10.1002/ajb2.16294","DOIUrl":null,"url":null,"abstract":"<p>Plants rely on adjustments in growth and development to respond to environmental stimuli. Developmental transitions, including germination, vegetative phase change, reproductive transition, and senescence, modify the growth patterns of plants and their requirements for survival. Consequently, the timing of developmental transitions and the developmental stage at which a plant encounters environmental stress hold significant implications for the performance of individuals, population dynamics, and community dynamics. If developmental phases, and the timing of transitions between them, are key to plant success in fluctuating environments, then understanding ontogenetic changes in plant environmental interactions is necessary to predict how plants will react to environmental stress and novel environments. Geneticists and molecular biologists have discovered many mechanisms governing developmental transitions, while developmental biologists have studied how plant form changes across ontogeny and ecologists have studied how plant form alters organismal interactions. However, there has been insufficient integration of these fields of study, hindering a comprehensive understanding of how plant development contributes to environmental adaptation and acclimation.</p><p>Links among plant development, plant fitness, and ecosystem functioning lead us to hypothesize that selection on the timing of developmental transitions to align stress-tolerant developmental phases with periods of environmental stress is more prevalent than currently understood. Phase-specific differences in environmental tolerance could contribute to genotypic and species-specific adaptations to climate. As depicted in Figure 1, two genotypes (or species) labeled “A” and “B” exhibit different patterns of phase-specific stress tolerance throughout ontogeny. Depending on the environment (Env. 1 or 2) they inhabit, they will experience harsh environmental conditions at different times. Genotypes (or species) that align stress-tolerant developmental phases with periods of harsh conditions (e.g., genotype “A” in Env. 1) are thus expected to have increased fitness.</p><p>This example illustrates that the alignment between stress-tolerant developmental phases and harsh environmental conditions can arise through changes in developmental timing or ontogenetic alterations in plant growth and physiology. Despite considerable progress in our understanding of relationships between ontogeny and environmental response that support this hypothesis, knowledge of when developmental transitions occur and the extent of genetic variation in the timing of these transitions and phase-specific traits remains limited.</p><p>As plants transition between developmental phases, their tolerance and response to environmental conditions changes (Figure 1). Plants in different developmental phases possess distinct organs (i.e., juvenile or adult leaves, flowers), access to resources, and physiological characteristics, which contribute to ontogenetic changes in environmental interactions. For example, during the transition from juvenile to adult vegetative phases, many plants produce different types of leaves that lead to shifts in carbon economics and suitability for different light environments. Juvenile leaves of multiple species are less costly to produce and have morphology and photosynthetic physiology better suited to low-light conditions, whereas adult leaves have a longer lifespan and have a morphology and physiology more profitable in high-light environments (Lawrence et al., <span>2022</span>).</p><p>Furthermore, plants in different developmental phases possess unique genome-wide expression responses to environmental stimuli that control physiological responses. For example, during vegetative phase change, age-dependent increases in the expression of SQUAMOSA PROMOTER BINDING-LIKE (SPL) genes activate defense signaling genes, enhancing disease resistance during the adult vegetative phase in <i>Arabidopsis thaliana</i> (L.) Heynh. [Brassicaceae] (Hu et al., <span>2023</span>). Additionally, there are phase-specific differences in gene expression responses to the environment, as observed in pre- and post-flowering <i>Sorghum bicolor</i> (L.) Moench [Poaceae] plants exposed to drought (Varoquaux et al., <span>2019</span>). Developmental stage-dependent environmental responses can be so significant that different species at the same developmental stage exhibit more similar responses to environment than conspecifics at different stages (Parrish and Bazzaz, <span>1985</span>). Although many published studies likely capture patterns of ontogenetic variation, failure to explicitly assess developmental stage may confound our ability to understand how plants interact with their environment across their lifespan, leaving a critical need for more research specifically focusing on phase-specific responses to environmental stress.</p><p>The timing of plant developmental transitions is intricately responsive to environmental fluctuations (Bäurle and Dean, <span>2006</span>). These plastic developmental responses are one of the most obvious plant responses to environment, noticeable even to casual observers. The most visually obvious transitions like flowering and senescence have long garnered the attention of humans, particularly in systems we depend on (e.g., crops) (Primack et al., <span>2009</span>). Interactions between developmental timing and environment not only alter the duration of each developmental phase, but also determine the stage at which a plant experiences specific stresses. For instance, some plants accelerate flowering in response to drought, thereby shortening their growing season and escaping end-of-season droughts (Ludlow, <span>1989</span>; Kooyers, <span>2015</span>).</p><p>Environmental impacts on developmental timing extend across multiple generations, increasing the likelihood of impacts on plant performance. For example, in <i>A. thaliana</i>, the flowering time of one generation influences the germination timing of the next, and vice versa (Burghardt et al., <span>2015</span>). Additionally, temperature during seed set affects seed dormancy, such that, seeds set in autumn tend to exhibit stronger dormancy than those set earlier in the growing season. This is particularly evident in genotypes adapted to warmer climates, where mild winter temperatures fail to maintain primary dormancy throughout the winter (Iwasaki et al., <span>2022</span>). While many studies have focused on environmental impacts on the timing of developmental transitions, there remains much unknown, especially regarding how multiple environmental factors and transitions interact and how complex fluctuations (e.g., precipitation in semiarid environments) influence developmental timing. Additionally, certain understudied developmental transitions, like the juvenile-to-adult vegetative transition known as vegetative phase change, continue to pose a significant knowledge gap that warrants further investigation.</p><p>The timing of developmental transitions and ontogenetic differences in environmental response contribute to a plant's fitness and survival across ecosystems, affecting a population's persistence under current and novel environmental conditions. Evolution of phenology, especially leaf out and flowering time, are possibly the most well-known mechanisms of local adaptation in plants (Alberto et al., <span>2013</span>). Studies show both genetic and plastic shifts in developmental timing within populations and communities in response to different environments, highlighting how such shifts affect individual performance across environments. An example of how selection of developmental transitions contribute to local adaptation is flowering time in <i>A. thaliana</i>, where researchers have found selection for either early or late flowering depending on environmental conditions (Korves et al., <span>2007</span>). Furthermore, <i>A. thaliana</i> has apparently adapted to climates across its geographic range using strategies that cause different genotypes to germinate and flower at different times of the year to match when local conditions are favorable for these developmental phases (Exposito-Alonso, <span>2020</span>; Martínez-Berdeja et al., <span>2020</span>).</p><p>Developmental transitions can alter selection and affect population, community, and ecosystem dynamics. For example, during early development, fast-growing “ruderal” strategies are often selected for and as plants age, more competitive and stress-tolerant strategies prevail (Dayrell et al., <span>2018</span>). Thus, plants in different developmental phases within the same ecosystem can deploy different growth strategies and changes in environmental conditions likely affect their performance in distinct ways. For instance, under water-stressed conditions, young <i>Artemisia californica</i> Less. [Asteraceae] plants maintain higher photosynthetic rates compared to older individuals however, these younger plants show greater drought sensitivity in other traits like stomatal conductance, whereas older plants are largely unaffected (Funk et al., <span>2021</span>).</p><p>These differences in growth strategies and resource requirements likely contribute to ontogenetic changes in interactions with neighboring plants (Niinemets, <span>2004</span>; Lasky et al., <span>2015</span>). For instance, <i>Dioon sonorense</i> (De Luca, Sabato & Vázq.Torres) Chemnick, T.Greg. & Salas-Morales [Zamiaceae] seedlings are more shade tolerant than individuals at later developmental stages. This leads to increases in interspecific competition with neighbors as plants age, because the canopy cover from neighboring species shifts from facilitating seedling survival to creating a light-limited environment for adults (Álvarez-Yépiz et al., <span>2014</span>). Ontogenetic changes in plant chemical and physical defense strategies alter competition with the non-plant community (e.g., herbivores) across development as well (Kariñho-Betancourt et al., <span>2015</span>). As the timing of developmental transitions shifts in response to the environment, the developmental phase in which species interact with one another, and the duration of these interactions, are also likely to change, with consequences for individual performance (Yang and Rudolf, <span>2010</span>). Due to the significance of how developmental transitions and phase-specific differences in environmental response contribute to a plant's fitness and survival across environments, more studies are needed to improve our understanding of the ecological implications of these responses.</p><p>Advancing knowledge of ontogenetic changes in ecophysiology and environmental response may yield better predictions of how plants cope with novel environments and shed light on potential adaptation in nature and breeding programs. As development represents a primary mechanism for sessile organisms to respond to environmental changes, we encourage researchers to intentionally consider plant developmental stages and transitions when designing experiments and constructing ecological and evolutionary models. Future studies that integrate molecular, organismal, and ecological research are needed to understand how the interactions among development, environment, and physiology contribute to plant fitness and ecosystem functioning, and how these interactions could provide insights for future agricultural and ecosystem management strategies.</p><p>E.H.L.-P. and J.R.L. both contributed to the conceptualization, writing, and revising of the manuscript.</p>","PeriodicalId":7691,"journal":{"name":"American Journal of Botany","volume":"111 3","pages":""},"PeriodicalIF":2.4000,"publicationDate":"2024-02-21","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1002/ajb2.16294","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"American Journal of Botany","FirstCategoryId":"99","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1002/ajb2.16294","RegionNum":2,"RegionCategory":"生物学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q2","JCRName":"PLANT SCIENCES","Score":null,"Total":0}
引用次数: 0
Abstract
Plants rely on adjustments in growth and development to respond to environmental stimuli. Developmental transitions, including germination, vegetative phase change, reproductive transition, and senescence, modify the growth patterns of plants and their requirements for survival. Consequently, the timing of developmental transitions and the developmental stage at which a plant encounters environmental stress hold significant implications for the performance of individuals, population dynamics, and community dynamics. If developmental phases, and the timing of transitions between them, are key to plant success in fluctuating environments, then understanding ontogenetic changes in plant environmental interactions is necessary to predict how plants will react to environmental stress and novel environments. Geneticists and molecular biologists have discovered many mechanisms governing developmental transitions, while developmental biologists have studied how plant form changes across ontogeny and ecologists have studied how plant form alters organismal interactions. However, there has been insufficient integration of these fields of study, hindering a comprehensive understanding of how plant development contributes to environmental adaptation and acclimation.
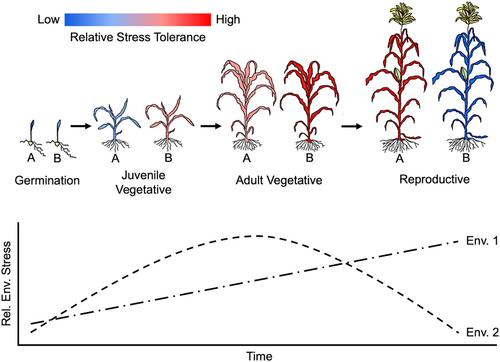
Links among plant development, plant fitness, and ecosystem functioning lead us to hypothesize that selection on the timing of developmental transitions to align stress-tolerant developmental phases with periods of environmental stress is more prevalent than currently understood. Phase-specific differences in environmental tolerance could contribute to genotypic and species-specific adaptations to climate. As depicted in Figure 1, two genotypes (or species) labeled “A” and “B” exhibit different patterns of phase-specific stress tolerance throughout ontogeny. Depending on the environment (Env. 1 or 2) they inhabit, they will experience harsh environmental conditions at different times. Genotypes (or species) that align stress-tolerant developmental phases with periods of harsh conditions (e.g., genotype “A” in Env. 1) are thus expected to have increased fitness.
This example illustrates that the alignment between stress-tolerant developmental phases and harsh environmental conditions can arise through changes in developmental timing or ontogenetic alterations in plant growth and physiology. Despite considerable progress in our understanding of relationships between ontogeny and environmental response that support this hypothesis, knowledge of when developmental transitions occur and the extent of genetic variation in the timing of these transitions and phase-specific traits remains limited.
As plants transition between developmental phases, their tolerance and response to environmental conditions changes (Figure 1). Plants in different developmental phases possess distinct organs (i.e., juvenile or adult leaves, flowers), access to resources, and physiological characteristics, which contribute to ontogenetic changes in environmental interactions. For example, during the transition from juvenile to adult vegetative phases, many plants produce different types of leaves that lead to shifts in carbon economics and suitability for different light environments. Juvenile leaves of multiple species are less costly to produce and have morphology and photosynthetic physiology better suited to low-light conditions, whereas adult leaves have a longer lifespan and have a morphology and physiology more profitable in high-light environments (Lawrence et al., 2022).
Furthermore, plants in different developmental phases possess unique genome-wide expression responses to environmental stimuli that control physiological responses. For example, during vegetative phase change, age-dependent increases in the expression of SQUAMOSA PROMOTER BINDING-LIKE (SPL) genes activate defense signaling genes, enhancing disease resistance during the adult vegetative phase in Arabidopsis thaliana (L.) Heynh. [Brassicaceae] (Hu et al., 2023). Additionally, there are phase-specific differences in gene expression responses to the environment, as observed in pre- and post-flowering Sorghum bicolor (L.) Moench [Poaceae] plants exposed to drought (Varoquaux et al., 2019). Developmental stage-dependent environmental responses can be so significant that different species at the same developmental stage exhibit more similar responses to environment than conspecifics at different stages (Parrish and Bazzaz, 1985). Although many published studies likely capture patterns of ontogenetic variation, failure to explicitly assess developmental stage may confound our ability to understand how plants interact with their environment across their lifespan, leaving a critical need for more research specifically focusing on phase-specific responses to environmental stress.
The timing of plant developmental transitions is intricately responsive to environmental fluctuations (Bäurle and Dean, 2006). These plastic developmental responses are one of the most obvious plant responses to environment, noticeable even to casual observers. The most visually obvious transitions like flowering and senescence have long garnered the attention of humans, particularly in systems we depend on (e.g., crops) (Primack et al., 2009). Interactions between developmental timing and environment not only alter the duration of each developmental phase, but also determine the stage at which a plant experiences specific stresses. For instance, some plants accelerate flowering in response to drought, thereby shortening their growing season and escaping end-of-season droughts (Ludlow, 1989; Kooyers, 2015).
Environmental impacts on developmental timing extend across multiple generations, increasing the likelihood of impacts on plant performance. For example, in A. thaliana, the flowering time of one generation influences the germination timing of the next, and vice versa (Burghardt et al., 2015). Additionally, temperature during seed set affects seed dormancy, such that, seeds set in autumn tend to exhibit stronger dormancy than those set earlier in the growing season. This is particularly evident in genotypes adapted to warmer climates, where mild winter temperatures fail to maintain primary dormancy throughout the winter (Iwasaki et al., 2022). While many studies have focused on environmental impacts on the timing of developmental transitions, there remains much unknown, especially regarding how multiple environmental factors and transitions interact and how complex fluctuations (e.g., precipitation in semiarid environments) influence developmental timing. Additionally, certain understudied developmental transitions, like the juvenile-to-adult vegetative transition known as vegetative phase change, continue to pose a significant knowledge gap that warrants further investigation.
The timing of developmental transitions and ontogenetic differences in environmental response contribute to a plant's fitness and survival across ecosystems, affecting a population's persistence under current and novel environmental conditions. Evolution of phenology, especially leaf out and flowering time, are possibly the most well-known mechanisms of local adaptation in plants (Alberto et al., 2013). Studies show both genetic and plastic shifts in developmental timing within populations and communities in response to different environments, highlighting how such shifts affect individual performance across environments. An example of how selection of developmental transitions contribute to local adaptation is flowering time in A. thaliana, where researchers have found selection for either early or late flowering depending on environmental conditions (Korves et al., 2007). Furthermore, A. thaliana has apparently adapted to climates across its geographic range using strategies that cause different genotypes to germinate and flower at different times of the year to match when local conditions are favorable for these developmental phases (Exposito-Alonso, 2020; Martínez-Berdeja et al., 2020).
Developmental transitions can alter selection and affect population, community, and ecosystem dynamics. For example, during early development, fast-growing “ruderal” strategies are often selected for and as plants age, more competitive and stress-tolerant strategies prevail (Dayrell et al., 2018). Thus, plants in different developmental phases within the same ecosystem can deploy different growth strategies and changes in environmental conditions likely affect their performance in distinct ways. For instance, under water-stressed conditions, young Artemisia californica Less. [Asteraceae] plants maintain higher photosynthetic rates compared to older individuals however, these younger plants show greater drought sensitivity in other traits like stomatal conductance, whereas older plants are largely unaffected (Funk et al., 2021).
These differences in growth strategies and resource requirements likely contribute to ontogenetic changes in interactions with neighboring plants (Niinemets, 2004; Lasky et al., 2015). For instance, Dioon sonorense (De Luca, Sabato & Vázq.Torres) Chemnick, T.Greg. & Salas-Morales [Zamiaceae] seedlings are more shade tolerant than individuals at later developmental stages. This leads to increases in interspecific competition with neighbors as plants age, because the canopy cover from neighboring species shifts from facilitating seedling survival to creating a light-limited environment for adults (Álvarez-Yépiz et al., 2014). Ontogenetic changes in plant chemical and physical defense strategies alter competition with the non-plant community (e.g., herbivores) across development as well (Kariñho-Betancourt et al., 2015). As the timing of developmental transitions shifts in response to the environment, the developmental phase in which species interact with one another, and the duration of these interactions, are also likely to change, with consequences for individual performance (Yang and Rudolf, 2010). Due to the significance of how developmental transitions and phase-specific differences in environmental response contribute to a plant's fitness and survival across environments, more studies are needed to improve our understanding of the ecological implications of these responses.
Advancing knowledge of ontogenetic changes in ecophysiology and environmental response may yield better predictions of how plants cope with novel environments and shed light on potential adaptation in nature and breeding programs. As development represents a primary mechanism for sessile organisms to respond to environmental changes, we encourage researchers to intentionally consider plant developmental stages and transitions when designing experiments and constructing ecological and evolutionary models. Future studies that integrate molecular, organismal, and ecological research are needed to understand how the interactions among development, environment, and physiology contribute to plant fitness and ecosystem functioning, and how these interactions could provide insights for future agricultural and ecosystem management strategies.
E.H.L.-P. and J.R.L. both contributed to the conceptualization, writing, and revising of the manuscript.
期刊介绍:
The American Journal of Botany (AJB), the flagship journal of the Botanical Society of America (BSA), publishes peer-reviewed, innovative, significant research of interest to a wide audience of plant scientists in all areas of plant biology (structure, function, development, diversity, genetics, evolution, systematics), all levels of organization (molecular to ecosystem), and all plant groups and allied organisms (cyanobacteria, algae, fungi, and lichens). AJB requires authors to frame their research questions and discuss their results in terms of major questions of plant biology. In general, papers that are too narrowly focused, purely descriptive, natural history, broad surveys, or that contain only preliminary data will not be considered.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


