{"title":"Stored alcohol and fatty acid intermediates and the biosynthesis of sex pheromone aldehyde in the moth Chloridea virescens.","authors":"Stephen P Foster, Karin G Anderson","doi":"10.1007/s10886-024-01478-x","DOIUrl":null,"url":null,"abstract":"<p><p>In most species of moths, the female produces and releases a volatile sex pheromone from a specific gland to attract a mate. Biosynthesis of the most common type of moth sex pheromone component (Type 1) involves de novo synthesis of hexadecanoate (16:Acyl), followed by modification to various fatty acyl intermediates, then reduction to a primary alcohol, which may be acetylated or oxidized to produce an acetate ester or aldehyde, respectively. Our previous work on the moth Chloridea virescens (Noctuidae) showed that females produce 90% of the major pheromone component, (Z)-11-hexadecenal (Z11-16:Ald), via a direct and rapid route of de novo biosynthesis with highly labile intermediates, and ca. 10% from an indirect route that likely mobilizes a pre-synthesized 16-carbon skeleton, possibly, (Z)-11-hexadecenoate (Z11-16:Acyl) or hexadecanoate (16:Acyl). In this paper, we use stable isotope tracer/tracee techniques to study the dynamics of the precursor alcohol (Z)-11-hexadecenol (Z11-16:OH) and stores of Z11-16:Acyl and 16:Acyl to determine their roles in biosynthesis of Z11-16:Ald. We found: (i) that intracellular Z11-16:OH is synthesized at roughly the same rate as Z11-16:Ald, indicating that translocation and oxidation of this moiety does not rate limit biosynthesis of Z11-16:Ald, (ii) intracellular Z11-16:OH consists of two pools, a highly labile one rapidly translocated out of the cell and converted to Z11-16:Ald, and a less labile one that mostly remains in gland cells, (iii) during pheromone biosynthesis, net stores of Z11-16:Acyl increase, suggesting it is not the source of Z11-16:Ald produced by the indirect route, and (iv) no evidence for the gland synthesizing stored 16:Acyl prior to (up to 2 days before eclosion), or after, synthesis of pheromone commenced, suggesting the bulk of this stored moiety is synthesized elsewhere and transported to the gland prior to gland maturation. Thus, the pheromone gland of C. virescens produces very little stored fat over its functional lifetime, being optimized to produce sex pheromone.</p>","PeriodicalId":2,"journal":{"name":"ACS Applied Bio Materials","volume":null,"pages":null},"PeriodicalIF":4.6000,"publicationDate":"2024-04-01","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://www.ncbi.nlm.nih.gov/pmc/articles/PMC11043202/pdf/","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"ACS Applied Bio Materials","FirstCategoryId":"93","ListUrlMain":"https://doi.org/10.1007/s10886-024-01478-x","RegionNum":0,"RegionCategory":null,"ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"2024/2/20 0:00:00","PubModel":"Epub","JCR":"Q2","JCRName":"MATERIALS SCIENCE, BIOMATERIALS","Score":null,"Total":0}
引用次数: 0
Abstract
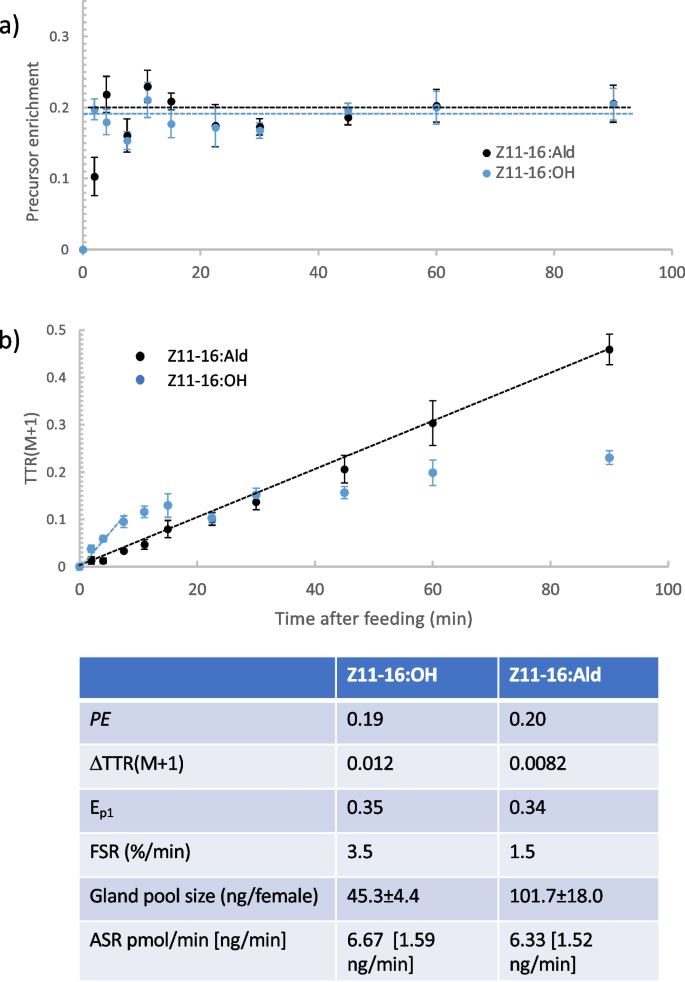
In most species of moths, the female produces and releases a volatile sex pheromone from a specific gland to attract a mate. Biosynthesis of the most common type of moth sex pheromone component (Type 1) involves de novo synthesis of hexadecanoate (16:Acyl), followed by modification to various fatty acyl intermediates, then reduction to a primary alcohol, which may be acetylated or oxidized to produce an acetate ester or aldehyde, respectively. Our previous work on the moth Chloridea virescens (Noctuidae) showed that females produce 90% of the major pheromone component, (Z)-11-hexadecenal (Z11-16:Ald), via a direct and rapid route of de novo biosynthesis with highly labile intermediates, and ca. 10% from an indirect route that likely mobilizes a pre-synthesized 16-carbon skeleton, possibly, (Z)-11-hexadecenoate (Z11-16:Acyl) or hexadecanoate (16:Acyl). In this paper, we use stable isotope tracer/tracee techniques to study the dynamics of the precursor alcohol (Z)-11-hexadecenol (Z11-16:OH) and stores of Z11-16:Acyl and 16:Acyl to determine their roles in biosynthesis of Z11-16:Ald. We found: (i) that intracellular Z11-16:OH is synthesized at roughly the same rate as Z11-16:Ald, indicating that translocation and oxidation of this moiety does not rate limit biosynthesis of Z11-16:Ald, (ii) intracellular Z11-16:OH consists of two pools, a highly labile one rapidly translocated out of the cell and converted to Z11-16:Ald, and a less labile one that mostly remains in gland cells, (iii) during pheromone biosynthesis, net stores of Z11-16:Acyl increase, suggesting it is not the source of Z11-16:Ald produced by the indirect route, and (iv) no evidence for the gland synthesizing stored 16:Acyl prior to (up to 2 days before eclosion), or after, synthesis of pheromone commenced, suggesting the bulk of this stored moiety is synthesized elsewhere and transported to the gland prior to gland maturation. Thus, the pheromone gland of C. virescens produces very little stored fat over its functional lifetime, being optimized to produce sex pheromone.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


