Song fragments recorded on a tagged juvenile humpback whale (Megaptera novaeangliae) on a winter feeding ground at the mouth of the Chesapeake Bay, Virginia, USA
Dana L. Adcock, Jeanne M. Shearer, Heather J. Foley, Zach T. Swaim, Andrew J. Read
{"title":"Song fragments recorded on a tagged juvenile humpback whale (Megaptera novaeangliae) on a winter feeding ground at the mouth of the Chesapeake Bay, Virginia, USA","authors":"Dana L. Adcock, Jeanne M. Shearer, Heather J. Foley, Zach T. Swaim, Andrew J. Read","doi":"10.1111/mms.13110","DOIUrl":null,"url":null,"abstract":"<p>Humpback whales produce a variety of sound types, including vocalizations termed “songs” because they consist of repeated, fixed patterns of sounds, similar to bird songs (Payne et al., <span>1971</span>). Humpback songs consist of themes, phrases, units, and subunits (Payne et al., <span>1971</span>). In most themes, phrases are repeated before the whale moves on to the next theme, with a complete song lasting 7–30 min (Payne et al., <span>1971</span>). Singers remain relatively stationary throughout the duration of their display (Helweg et al., <span>1992</span>).</p><p>Humpback songs are produced most frequently on breeding grounds where they are used as a reproductive display by adult males (Clark & Clapham <span>2004</span>; Eriksen et al., <span>2005</span>; Vu et al., <span>2012</span>) and contribute to male reproductive success (Darling & Bérubé, <span>2001</span>; Helweg et al., <span>1992</span>; Tyack, <span>1983</span>; Tyack & Clark, <span>2000</span>; Tyack & Whitehead, <span>1983</span>). The complexity of song may be attractive to females and thus serve a function in sexual selection (Tyack, <span>1983</span>). Various aspects of the components of a song, including novelty, may serve as indicators of fitness in male whales (Helweg et al., <span>1992</span>). Whales incorporate components of song from other populations, suggesting that novelty contributes to the evolution of humpback whale song (Noad et al., <span>2000</span>) and songs change continuously in a directional pattern driven by cultural transmission (Eriksen et al., <span>2005</span>).</p><p>Song has also been recorded on migration routes and on high-latitude feeding grounds (Martin et al., <span>2021</span>; Mattila et al., <span>1987</span>; Vu et al., <span>2012</span>). For example, Mattila et al. (<span>1987</span>) recorded songs on Stellwagen Bank, Massachusetts, a high-latitude summer feeding ground, in the autumn of 1983–1984. Songs were also recorded on the same feeding ground in the summer and spring of 2003 and year-round in 2006 and 2008 (Clark & Clapham, <span>2004</span>; Vu et al., <span>2012</span>). In year-round recordings, song occurrence and duration increased in late autumn through December as males approached southward migration periods (Vu et al., <span>2012</span>). Whales from the Australian and New Caledonian populations sing on high-latitude Antarctic summer feeding grounds (Garland et al., <span>2013</span>) and males sing on Southeast Alaskan summer feeding grounds (McSweeney et al., <span>1989</span>). Recently, humpbacks have been recorded singing over several months in the winter and spring on Norwegian feeding grounds (Martin et al., <span>2021</span>). Song has been recorded on migration routes between breeding grounds in the Caribbean and feeding grounds in the North Atlantic and in the central New Zealand migratory corridor (Charif et al., <span>2001</span>; Vu et al., <span>2012</span>; Warren et al., <span>2020</span>). Song has also been recorded in the New York Bight (Zeh et al., <span>2021</span>). Thus, singing behavior is not confined to low-latitude breeding grounds. In addition to song, humpback whales also use other types of vocalizations and behaviors to communicate while foraging on high-latitude feeding grounds (Parks et al., <span>2014</span>).</p><p>The North Atlantic population of humpback whales migrate from high-latitude summer feeding grounds in the northeastern U.S., Canada, Iceland, and Greenland to winter breeding grounds in the Caribbean (Katona et al., <span>1983</span>; Leatherwood et al., <span>1976</span>; Martin et al., <span>1984</span>; Swingle et al., <span>1993</span>). However, some humpback whales, primarily juveniles, do not complete the migration to breeding grounds and, instead, remain on nearshore mid-latitude feeding grounds along the mid-Atlantic coast, where they feed during winter (Swingle et al., <span>1993</span>). We recorded song-like fragments on a digital acoustic tag (DTAG) attached to a humpback whale in the nearshore waters of Virginia Beach, Virginia; to our knowledge this is the first humpback song recorded on this mid-latitude winter feeding ground.</p><p>This tagged whale was one of 11 animals we equipped with version 3 DTAGs (Johnson & Tyack, <span>2003</span>) at the mouth of the Chesapeake Bay between 2019 and 2021 (Figure 1; Shearer et al., <span>2021</span>). These tags include two hydrophones sampling at 120 or 240 kHz to record sound and triaxial accelerometers and magnetometers sampling at 250 Hz to record movement. We deployed the tag using a 7-m carbon-fiber pole. The tag emits a VHF signal detectable when the tagged whale is at the surface. We conducted a focal follow during the entire 5-hr deployment, recording the tagged whale's location and behavior. No other whales were sighted during the focal follow of the tagged whale (5 hr). Once the tag released from the whale, it floated to the surface, where we located it using the VHF signal.</p><p>We calibrated the tag using the DTAG toolbox (https://animaltags.org) and custom-written scripts, computing the animal's pitch, roll, and heading using the orientation of the animal during surfacings (Johnson, <span>2011</span>). Using MATLAB 2016b (The Mathworks, Inc.; https://www.mathworks.com/), we high-pass filtered the data at 100 Hz to remove flow noise. We used Raven, a bioacoustics software, for waveform and spectrogram analysis (K. Lisa Yang Center for Conservation Bioacoustics at the Cornell Lab of Ornithology, <span>2022</span>). We adjusted the spectrogram to a fast Fourier transform window size of 512. We set the spectrogram viewing window at 0–8 kHz. One analyst (DLA) listened to audio while analyzing the spectrogram and manually identified 11 subjective call types. We used MATLAB 2016b (The Mathworks, Inc.) to calculate the following acoustic parameters: duration, noise level, signal level, SNR, peak frequency, lower 10 dB re: 1 μPa frequency, and upper 10 dB re: 1 μPa frequency (Table 1). Signal and noise levels were estimated as the root mean square (RMS) amplitude, corrected for a nominal tag sensitivity (−175 dB re: 1 V μPa<sup>−1</sup>; Silva et al., <span>2017</span>). Noise levels were taken over 0.2 s prior to the unit, manually checking that there were no other vocalizations in that window. This particular tag did not have a sensitivity calibration, so signal levels may not be exact, but the ratio between signal and noise level is not affected by this calibration. We assigned call types to descriptions in the literature based on the spectrogram image, duration, frequency, and presence of repetition (Dunlop et al., <span>2007</span>; Fournet et al., <span>2015</span>). We excluded vocalizations with a signal-to-noise ratio (SNR) under 10 dB re: 1 μPa to ensure the calls were produced near the DTAG (Stimpert et al., <span>2020</span>). While there were no other whales visible during the deployment, it cannot be confirmed that the vocalizations are from the tagged whale. The SNR threshold ensures that the calls were produced close to the tag. The signal level for each call type remained relatively consistent throughout the recording.</p><p>We marked the start and end times of each vocalization by eye using the sound levels in the waveform and the signal markings in the spectrogram (Rekdahl et al., <span>2013</span>). After we identified the type of each call, we visually analyzed the selection tables in Microsoft Excel to look for patterns in unit types. We grouped varying numbers of repeating units with the same general pattern into phrases. We color-coded the phrases and then identified themes by marking distinct repetitions of phrases. For example, unit types K and H formed variations of “KH” which was repeated six times over the course of 48 s. We then separated different themes by noting distinct transitions from one phrase repetition to another with notable changes in structure and unit types used. We defined a song fragment as one subphrase of two or more themes (not repeated), and songs as three or more themes repeated at least once (Kowarski et al., <span>2019</span>). We categorized themes into song fragments by tracking the patterned repetition of themes alongside the time duration. Two analysts (D.L.A. and J.M.S.) performed this phrase, theme, and song fragment analysis separately, then compared and identified matching findings. We only included phrases that both analysts agreed on.</p><p>The tagged whale measured 9.0 m in length (measured via drone photogrammetry by our colleagues at HDR, Inc.; see Aschettino et al., <span>2022</span>), which would classify it as a juvenile or subadult. Unfortunately, we were unable to collect a biopsy sample to determine the sex of the whale. During the 5-hr tag recording, the whale vocalized for 3 h and 10 min, stopped for 30 min, and then continued for 9 min more until the tag detached. The whale did not vocalize for the first hour. During this vocally active period, we identified 11 distinct call types (Figure 2, Table 2). Call type B appears to be a nonsong call and has been hypothesized to be a form of social communication to other whales (Dunlop et al., <span>2007</span>, <span>2008</span>). The other call types are units found in song structure (Dunlop et al., <span>2007</span>; Fournet et al., <span>2015</span>). No vocalizations overlapped in time.</p><p>We identified and color-coded seven total phrases (Figure 3). There was clear patterning in both unit types and phrases, which form themes. For example, a theme of phrases 5, 1, 2, and 3 appears multiple times (Figure 3). Phrases 6 and 4 and phrases 7 and 4 also form themes (Figure 3). Some phrases repeated consecutively without interjection of other phrases or nonsong calls (Figure 3).</p><p>During a period of 33 min, while it produced 9 of the 10 song unit types, the tagged whale positioned itself head down with a pitch angle between −20 and − 60 degrees, a common orientation observed in singing adult males on breeding grounds (Au et al., <span>2006</span>; Herman et al., <span>2013</span>). During this time, the tagged whale's dive depths ranged from 10 to 14 m. Both the focal follow and accelerometry data from the tag showed relatively little movement during this period as the tagged whale drifted passively during these vocalizations.</p><p>We conclude that the clear, patterned vocal themes provide evidence of humpback song fragments produced on this mid-latitude, winter feeding ground (Swingle et al., <span>1993</span>). The units created distinct phrases, which formed themes, and the themes themselves were organized. These patterns indicate the presence of song fragments as defined in the literature (Kowarski et al., <span>2019</span>). While there was some variation, the identified themes show consistently repeated phrases. We hypothesize that the tagged whale was a juvenile male practicing singing behavior. The purpose of nonsong calls between song fragments is still unclear and should be addressed by future research. Previous research on humpback song has largely focused on adults and little is known about the development of this behavior, nor how it is used on feeding grounds. Our observations help to fill this gap.</p><p>Vessel strikes are a major threat to humpback whales (Stepanuk et al., <span>2021</span>) and collisions are more common along the U.S. Atlantic Coast than anywhere else in North America (Jensen & Silber, <span>2003</span>). This population of humpback whales have been undergoing an Unusual Mortality Event since 2016, with 40% of deaths in the past year attributed to ship strikes and entanglement (Office of Protected Resources, <span>2023</span>). Juvenile humpback whales in the mid-Atlantic feed nearshore in New York and Virginia (Stepanuk et al., <span>2021</span>; Swingle et al., <span>1993</span>). For these individuals, incidences of ship strikes may be greater, since ship traffic is typically increased in these areas (Aschettino et al., <span>2020</span>; Shearer et al., <span>2021</span>). This whale was approximately 8–11 km from shore and in or just outside shipping lanes for the duration of the deployment. There were at least three ships with a draft between 12.3 and 12.8 m in the area, which would overlap with the depth of the whale. The depth of the area is approximately 9–12 m outside of the shipping lanes, with lanes dredged to about 25 m. The singing behavior we documented here may increase this whale's risk of being struck, as its passive, head-down orientation between 10 and 14 m increases its vulnerability to approaching ships. Here we document song produced on this winter, mid-latitude feeding ground, contributing to our understanding of juvenile humpback whale behavior. Future research should expand on reductions in communication space and increased ship-strike risk for developing singers.</p><p><b>Dana L. Adcock:</b> Data curation; formal analysis; software; visualization; writing – original draft; writing – review and editing. <b>Jeanne M. Shearer:</b> Conceptualization; formal analysis; funding acquisition; investigation; methodology; project administration; software; supervision; validation; visualization; writing – review and editing. <b>Heather J. Foley:</b> Investigation; writing – review and editing. <b>Zach T. Swaim:</b> Investigation; project administration; supervision. <b>Andrew J. Read:</b> Conceptualization; formal analysis; funding acquisition; methodology; project administration; software; supervision; validation; visualization; writing – review and editing.</p><p>The authors declare they have no conflicts of interest.</p>","PeriodicalId":18725,"journal":{"name":"Marine Mammal Science","volume":"40 3","pages":""},"PeriodicalIF":2.0000,"publicationDate":"2024-02-02","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1111/mms.13110","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"Marine Mammal Science","FirstCategoryId":"99","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1111/mms.13110","RegionNum":3,"RegionCategory":"生物学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q2","JCRName":"MARINE & FRESHWATER BIOLOGY","Score":null,"Total":0}
引用次数: 0
Abstract
Humpback whales produce a variety of sound types, including vocalizations termed “songs” because they consist of repeated, fixed patterns of sounds, similar to bird songs (Payne et al., 1971). Humpback songs consist of themes, phrases, units, and subunits (Payne et al., 1971). In most themes, phrases are repeated before the whale moves on to the next theme, with a complete song lasting 7–30 min (Payne et al., 1971). Singers remain relatively stationary throughout the duration of their display (Helweg et al., 1992).
Humpback songs are produced most frequently on breeding grounds where they are used as a reproductive display by adult males (Clark & Clapham 2004; Eriksen et al., 2005; Vu et al., 2012) and contribute to male reproductive success (Darling & Bérubé, 2001; Helweg et al., 1992; Tyack, 1983; Tyack & Clark, 2000; Tyack & Whitehead, 1983). The complexity of song may be attractive to females and thus serve a function in sexual selection (Tyack, 1983). Various aspects of the components of a song, including novelty, may serve as indicators of fitness in male whales (Helweg et al., 1992). Whales incorporate components of song from other populations, suggesting that novelty contributes to the evolution of humpback whale song (Noad et al., 2000) and songs change continuously in a directional pattern driven by cultural transmission (Eriksen et al., 2005).
Song has also been recorded on migration routes and on high-latitude feeding grounds (Martin et al., 2021; Mattila et al., 1987; Vu et al., 2012). For example, Mattila et al. (1987) recorded songs on Stellwagen Bank, Massachusetts, a high-latitude summer feeding ground, in the autumn of 1983–1984. Songs were also recorded on the same feeding ground in the summer and spring of 2003 and year-round in 2006 and 2008 (Clark & Clapham, 2004; Vu et al., 2012). In year-round recordings, song occurrence and duration increased in late autumn through December as males approached southward migration periods (Vu et al., 2012). Whales from the Australian and New Caledonian populations sing on high-latitude Antarctic summer feeding grounds (Garland et al., 2013) and males sing on Southeast Alaskan summer feeding grounds (McSweeney et al., 1989). Recently, humpbacks have been recorded singing over several months in the winter and spring on Norwegian feeding grounds (Martin et al., 2021). Song has been recorded on migration routes between breeding grounds in the Caribbean and feeding grounds in the North Atlantic and in the central New Zealand migratory corridor (Charif et al., 2001; Vu et al., 2012; Warren et al., 2020). Song has also been recorded in the New York Bight (Zeh et al., 2021). Thus, singing behavior is not confined to low-latitude breeding grounds. In addition to song, humpback whales also use other types of vocalizations and behaviors to communicate while foraging on high-latitude feeding grounds (Parks et al., 2014).
The North Atlantic population of humpback whales migrate from high-latitude summer feeding grounds in the northeastern U.S., Canada, Iceland, and Greenland to winter breeding grounds in the Caribbean (Katona et al., 1983; Leatherwood et al., 1976; Martin et al., 1984; Swingle et al., 1993). However, some humpback whales, primarily juveniles, do not complete the migration to breeding grounds and, instead, remain on nearshore mid-latitude feeding grounds along the mid-Atlantic coast, where they feed during winter (Swingle et al., 1993). We recorded song-like fragments on a digital acoustic tag (DTAG) attached to a humpback whale in the nearshore waters of Virginia Beach, Virginia; to our knowledge this is the first humpback song recorded on this mid-latitude winter feeding ground.
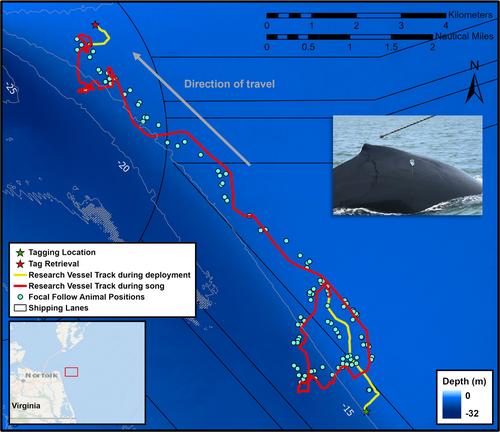
This tagged whale was one of 11 animals we equipped with version 3 DTAGs (Johnson & Tyack, 2003) at the mouth of the Chesapeake Bay between 2019 and 2021 (Figure 1; Shearer et al., 2021). These tags include two hydrophones sampling at 120 or 240 kHz to record sound and triaxial accelerometers and magnetometers sampling at 250 Hz to record movement. We deployed the tag using a 7-m carbon-fiber pole. The tag emits a VHF signal detectable when the tagged whale is at the surface. We conducted a focal follow during the entire 5-hr deployment, recording the tagged whale's location and behavior. No other whales were sighted during the focal follow of the tagged whale (5 hr). Once the tag released from the whale, it floated to the surface, where we located it using the VHF signal.
We calibrated the tag using the DTAG toolbox (https://animaltags.org) and custom-written scripts, computing the animal's pitch, roll, and heading using the orientation of the animal during surfacings (Johnson, 2011). Using MATLAB 2016b (The Mathworks, Inc.; https://www.mathworks.com/), we high-pass filtered the data at 100 Hz to remove flow noise. We used Raven, a bioacoustics software, for waveform and spectrogram analysis (K. Lisa Yang Center for Conservation Bioacoustics at the Cornell Lab of Ornithology, 2022). We adjusted the spectrogram to a fast Fourier transform window size of 512. We set the spectrogram viewing window at 0–8 kHz. One analyst (DLA) listened to audio while analyzing the spectrogram and manually identified 11 subjective call types. We used MATLAB 2016b (The Mathworks, Inc.) to calculate the following acoustic parameters: duration, noise level, signal level, SNR, peak frequency, lower 10 dB re: 1 μPa frequency, and upper 10 dB re: 1 μPa frequency (Table 1). Signal and noise levels were estimated as the root mean square (RMS) amplitude, corrected for a nominal tag sensitivity (−175 dB re: 1 V μPa−1; Silva et al., 2017). Noise levels were taken over 0.2 s prior to the unit, manually checking that there were no other vocalizations in that window. This particular tag did not have a sensitivity calibration, so signal levels may not be exact, but the ratio between signal and noise level is not affected by this calibration. We assigned call types to descriptions in the literature based on the spectrogram image, duration, frequency, and presence of repetition (Dunlop et al., 2007; Fournet et al., 2015). We excluded vocalizations with a signal-to-noise ratio (SNR) under 10 dB re: 1 μPa to ensure the calls were produced near the DTAG (Stimpert et al., 2020). While there were no other whales visible during the deployment, it cannot be confirmed that the vocalizations are from the tagged whale. The SNR threshold ensures that the calls were produced close to the tag. The signal level for each call type remained relatively consistent throughout the recording.
We marked the start and end times of each vocalization by eye using the sound levels in the waveform and the signal markings in the spectrogram (Rekdahl et al., 2013). After we identified the type of each call, we visually analyzed the selection tables in Microsoft Excel to look for patterns in unit types. We grouped varying numbers of repeating units with the same general pattern into phrases. We color-coded the phrases and then identified themes by marking distinct repetitions of phrases. For example, unit types K and H formed variations of “KH” which was repeated six times over the course of 48 s. We then separated different themes by noting distinct transitions from one phrase repetition to another with notable changes in structure and unit types used. We defined a song fragment as one subphrase of two or more themes (not repeated), and songs as three or more themes repeated at least once (Kowarski et al., 2019). We categorized themes into song fragments by tracking the patterned repetition of themes alongside the time duration. Two analysts (D.L.A. and J.M.S.) performed this phrase, theme, and song fragment analysis separately, then compared and identified matching findings. We only included phrases that both analysts agreed on.
The tagged whale measured 9.0 m in length (measured via drone photogrammetry by our colleagues at HDR, Inc.; see Aschettino et al., 2022), which would classify it as a juvenile or subadult. Unfortunately, we were unable to collect a biopsy sample to determine the sex of the whale. During the 5-hr tag recording, the whale vocalized for 3 h and 10 min, stopped for 30 min, and then continued for 9 min more until the tag detached. The whale did not vocalize for the first hour. During this vocally active period, we identified 11 distinct call types (Figure 2, Table 2). Call type B appears to be a nonsong call and has been hypothesized to be a form of social communication to other whales (Dunlop et al., 2007, 2008). The other call types are units found in song structure (Dunlop et al., 2007; Fournet et al., 2015). No vocalizations overlapped in time.
We identified and color-coded seven total phrases (Figure 3). There was clear patterning in both unit types and phrases, which form themes. For example, a theme of phrases 5, 1, 2, and 3 appears multiple times (Figure 3). Phrases 6 and 4 and phrases 7 and 4 also form themes (Figure 3). Some phrases repeated consecutively without interjection of other phrases or nonsong calls (Figure 3).
During a period of 33 min, while it produced 9 of the 10 song unit types, the tagged whale positioned itself head down with a pitch angle between −20 and − 60 degrees, a common orientation observed in singing adult males on breeding grounds (Au et al., 2006; Herman et al., 2013). During this time, the tagged whale's dive depths ranged from 10 to 14 m. Both the focal follow and accelerometry data from the tag showed relatively little movement during this period as the tagged whale drifted passively during these vocalizations.
We conclude that the clear, patterned vocal themes provide evidence of humpback song fragments produced on this mid-latitude, winter feeding ground (Swingle et al., 1993). The units created distinct phrases, which formed themes, and the themes themselves were organized. These patterns indicate the presence of song fragments as defined in the literature (Kowarski et al., 2019). While there was some variation, the identified themes show consistently repeated phrases. We hypothesize that the tagged whale was a juvenile male practicing singing behavior. The purpose of nonsong calls between song fragments is still unclear and should be addressed by future research. Previous research on humpback song has largely focused on adults and little is known about the development of this behavior, nor how it is used on feeding grounds. Our observations help to fill this gap.
Vessel strikes are a major threat to humpback whales (Stepanuk et al., 2021) and collisions are more common along the U.S. Atlantic Coast than anywhere else in North America (Jensen & Silber, 2003). This population of humpback whales have been undergoing an Unusual Mortality Event since 2016, with 40% of deaths in the past year attributed to ship strikes and entanglement (Office of Protected Resources, 2023). Juvenile humpback whales in the mid-Atlantic feed nearshore in New York and Virginia (Stepanuk et al., 2021; Swingle et al., 1993). For these individuals, incidences of ship strikes may be greater, since ship traffic is typically increased in these areas (Aschettino et al., 2020; Shearer et al., 2021). This whale was approximately 8–11 km from shore and in or just outside shipping lanes for the duration of the deployment. There were at least three ships with a draft between 12.3 and 12.8 m in the area, which would overlap with the depth of the whale. The depth of the area is approximately 9–12 m outside of the shipping lanes, with lanes dredged to about 25 m. The singing behavior we documented here may increase this whale's risk of being struck, as its passive, head-down orientation between 10 and 14 m increases its vulnerability to approaching ships. Here we document song produced on this winter, mid-latitude feeding ground, contributing to our understanding of juvenile humpback whale behavior. Future research should expand on reductions in communication space and increased ship-strike risk for developing singers.
Dana L. Adcock: Data curation; formal analysis; software; visualization; writing – original draft; writing – review and editing. Jeanne M. Shearer: Conceptualization; formal analysis; funding acquisition; investigation; methodology; project administration; software; supervision; validation; visualization; writing – review and editing. Heather J. Foley: Investigation; writing – review and editing. Zach T. Swaim: Investigation; project administration; supervision. Andrew J. Read: Conceptualization; formal analysis; funding acquisition; methodology; project administration; software; supervision; validation; visualization; writing – review and editing.
The authors declare they have no conflicts of interest.
期刊介绍:
Published for the Society for Marine Mammalogy, Marine Mammal Science is a source of significant new findings on marine mammals resulting from original research on their form and function, evolution, systematics, physiology, biochemistry, behavior, population biology, life history, genetics, ecology and conservation. The journal features both original and review articles, notes, opinions and letters. It serves as a vital resource for anyone studying marine mammals.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


