{"title":"Can nitrate-reducing ammonifiers increase nitrogen retention in soil and support ammonium-based cropping systems?","authors":"Sara Hallin, Aurélien Saghaï","doi":"10.1002/sae2.12073","DOIUrl":null,"url":null,"abstract":"<p>Crop production depends on input of nitrogen (N) but because N-use efficiency is low in current conventional cropping systems, farmers fertilize much more than the plants need. More than 50% of added fertilizer N is lost to the environment, mainly as nitrate and gaseous N, that is, dinitrogen, nitrous oxide (N<sub>2</sub>O), and ammonia (Lassaletta et al., <span>2014</span>). Apart from deteriorating water quality and negatively impacting biodiversity, a main concern is the emissions of the greenhouse gas N<sub>2</sub>O. Nitrous oxide exhibits a global warming potential approximately 300 times higher than that of CO<sub>2</sub>, and the N<sub>2</sub>O concentration in the atmosphere is increasing at an accelerating rate (Thompson et al., <span>2019</span>). Anthropogenic sources contribute ca. 45% to global N<sub>2</sub>O emissions, with direct and indirect emissions from N additions in agriculture accounting for ca. 50% (Tian et al., <span>2020</span>). The negative consequences of N fertilisation therefore make the global food system a key target to limit climate change (Clark et al., <span>2020</span>) and allow humanity to remain within a safe operating space of the Earth system.</p><p>A main challenge for sustainable agriculture is to increase N-use efficiency in cropping systems without compromising yields. One possibility is to improve the retention of soil N by increasing the time N stays in the form of ammonium, as ammonium adsorbs clay particles and soil organic matter. This can be done by using nitrification inhibitors that hinder the microbially mediated oxidation of ammonium to nitrate (Coskun et al., <span>2017</span>) or by supporting nitrate ammonification, an overlooked process in the N cycle in which nitrate is reduced via nitrite to ammonium (a process also known as dissimilatory nitrate reduction to ammonium [DNRA]). Similar to the competing process of denitrification, nitrate ammonification is performed by phylogenetically diverse microorganisms, which couple the oxidation of various electron donors, most often organic carbon compounds, to the reduction of nitrate under anoxic conditions. Nitrate ammonification creates a short-circuit in the N cycle, bypassing denitrification and N-fixation, and can thereby contribute to primary production (Figure 1). There is, however, a possible risk of ammonia volatilization in alkaline soils. By contrast, the reduction of nitrate to gaseous N oxides through denitrification always results in ecosystem N losses, with a substantial amount emitted as N<sub>2</sub>O. Thus, the predominant nitrate reduction pathway affects the fate of nitrate and may have major consequences for N-use efficiency in cropping systems and possibly also climate change.</p><p>In this commentary, we highlight challenges and key research questions that need to be addressed to be able to evaluate the promises of nitrate ammonification and the feasibility of exploiting this process in sustainable agriculture. These include (i) estimating the relative importance of ammonium production rates via ammonification versus mineralisation, (ii) determining the contribution of nitrate ammonifiers to N<sub>2</sub>O reduction and production, (iii) assessing the biotic and abiotic factors promoting nitrate ammonification over denitrification, and (iv) exploring the possibility of harnessing plant traits to promote nitrate ammonification and increase N-use efficiency in cropping systems.</p><p>Nitrate ammonification rates vary greatly within croplands and managed grasslands (Supporting Information: Table S1). However, it is uncertain how much N it contributes in relation to other sources of ammonium in soil. When comparing these rates to gross N mineralization rates, nitrate ammonification contributes between 0% and 50% (average about 6% and median about 1%; Supporting Information: Table S1) of the produced ammonium in croplands and grasslands, without accounting for the release of previously produced ammonium adsorbed to soil particles. For the comparison, we mainly selected studies based on soil incubation assays with <sup>15</sup>N isotopes combined with tracing models using numerical solutions, which allows simultaneous quantification and comparison of multiple N transformations (Rütting et al., <span>2011</span>). Although these studies suggest that nitrate ammonification could be relevant for sustainable agriculture, the estimates are based on a limited number of studies and soil types as well as different models and assumptions. Further, the concentrations of ammonium and nitrate after substrate addition may also affect the fate of nitrate, considering that nitrate ammonifiers are favoured under low nitrate levels (Saghaï et al., <span>2023</span>; van den Berg et al., <span>2016</span>). Additional work is thus needed to gain both a better quantitative understanding of soil N fluxes, also acknowledging sorption and release of ammonium across years in the crop rotation, and a more representative picture of the relative importance of ammonification versus mineralization for delivering ammonium in cropping systems.</p><p>One of the possible benefits of enhancing ammonification is the reduction of N<sub>2</sub>O emissions, as shown by the negative correlation between nitrate ammonification rates and N<sub>2</sub>O emissions at the ecosystem scale across terrestrial biomes (Cheng et al., <span>2022</span>). However, only unfertilised soils were included in that analysis, and there is limited information on this relationship in fertilised agricultural soils (Putz et al., <span>2018</span>). Although the nitrate ammonification pathway in itself should not result in N<sub>2</sub>O production, small amounts of N<sub>2</sub>O have been reported from nitrate ammonifier isolates grown under conditions favourable to nitrate ammonification, for example (Stremińska et al., <span>2012</span>), which has been attributed to detoxification of nitric oxide-mediated by the ammonia forming cytochrome c nitrite reductase, NrfA (Poock et al., <span>2002</span>). At the same time, several ammonifiers have also been shown to carry <i>nosZ</i>, coding for the N<sub>2</sub>O reductase, and it has even been discussed that they could play a role as N<sub>2</sub>O sinks (Hallin et al., <span>2018</span>). Nevertheless, a recent analysis of ca. 1100 nitrate ammonifier genomes showed that capacity for denitrification and/or nitric oxide reduction is quite common, with 45% carrying one or several denitrification genes and more often for N<sub>2</sub>O production than reduction (Saghaï et al., <span>2023</span>). This implies that nitrate ammonifiers play a more complex role in the N cycle than previously thought and underlines that both the community composition of ammonifiers and environmental conditions matter for soil N<sub>2</sub>O fluxes and the retention of N in agroecosystems.</p><p>A recent soil metagenome screening of the gene <i>nrfA</i>, encoding NrfA, revealed that there is a large untapped potential for nitrate ammonification in both croplands and the rhizosphere, only comparable to tropical moist forests (Saghaï et al., <span>2023</span>). A key issue is how to make the best use of this genetic potential while minimizing the potential for denitrification that typically dominates over that of nitrate ammonification in nearly all terrestrial biomes, especially in croplands (Saghaï et al., <span>2023</span>). Our understanding of the factors influencing the fate of nitrate in soils is still limited (Cheng et al., <span>2022</span>), although a few management practices leading to higher soil organic carbon and carbon-to-nitrate ratio, like the inclusion of short-term grasslands in crop rotations, straw incorporation and no-tillage, have been shown to increase nitrate ammonification rates or the relative importance of nitrate ammonification to denitrification at single locales (Putz et al., <span>2018</span>; Yuan, Liu, et al., <span>2022</span>; Table 1). However, practices aiming to increase carbon sequestration should also optimize N fertilization strategies, especially in soils with low carbon content (Saghaï et al., <span>2023</span>). Recent work has highlighted the potential of biochar with high electron shuttle capacity to support nitrate ammonification in rice paddies (Yuan, Wang, et al., <span>2022</span>), as this pathway requires more electrons to be transferred than denitrification (Tiedje et al., <span>1982</span>). Although it is more likely to achieve effective N retention in cropping systems if ammonium production and plant uptake are closely coupled, most studies comparing denitrification and ammonification are typically conducted in ‘bulk’ soil and knowledge of plant controls on nitrate ammonification as well as on the competition between the two processes in the root environment is lacking.</p><p>Exploiting plant–microbe interactions has been identified as a promising strategy to develop sustainable crops. Large efforts are devoted to identify relevant plant functional traits and their genetic determinants (Bergelson et al., <span>2021</span>), also for improving N cycling in agroecosystems (Abalos et al., <span>2019</span>). Plants modify the physical and chemical conditions in the rhizosphere in both time and space, affecting the two major factors regulating the microbial reduction of nitrate, that is, the availability of the main substrates (nitrate and easily accessible organic carbon) and the oxygen partial pressure. Thus, plants with high N uptake efficiency and high growth rates exacerbate the competition for mineral N (Moreau et al., <span>2015</span>). This could possibly create a more favourable environment for ammonification than for denitrification, as enzymes used in ammonification typically display a lower affinity for nitrate and nitrite (van den Berg et al., <span>2016</span>). Moreover, the quantity and quality of organic carbon affect not only denitrification rates and the abundance of denitrifying microorganisms (Henry et al., <span>2008</span>) but also the partitioning of nitrate between the two competing pathways (Carlson et al., <span>2020</span>), although this has not been demonstrated <i>in planta</i> with natural root exudates. With the rhizosphere being a hotspot for microbial activity, including N assimilation and transformations (Philippot et al., <span>2013</span>), understanding how plant traits can favour nitrate ammonification over denitrification and allow plants to take advantage of the ammonium produced by ammonifiers will be crucial for the selection and breeding of climate-smart crop varieties (Table 1).</p><p>Conservation of soil N as ammonium means transitioning from nitrate to ammonium-based agriculture (Subbarao & Searchinger, <span>2021</span>). Apart from decreasing N losses, this could also lead to higher crop yields if ammonium-to-nitrate ratios in soil are optimized. Indeed, a mix of the two forms of reactive N can improve crop yields compared with conditions with only nitrate (Wang et al., <span>2019</span>), likely by making use of the plant's separate systems for the assimilation of ammonium and nitrate and thereby increasing N uptake efficiency. Similarly, ammonium has been shown to decrease plant N uptake if nitrate levels are low (Kuppe & Postma, <span>2023</span>). Ammonium can also be toxic to plants, although responses to ammonium vary not only among crops but also among crop varieties. This can be exploited for selection and breeding for crops that profit from higher ammonium levels. Improving soil N retention by promoting nitrate ammonification should work hand in hand with other efforts to improve N-use efficiency based on further developing synthetic nitrification inhibitors and biological nitrification inhibition since all these approaches rely on more ammonium-based cropping systems.</p><p>Sara Hallin and Aurélien Saghaï conceived the study and acquired funding. Sara Hallin and Aurélien Saghaï compiled the data in Table S1. Sara Hallin wrote the original draft, and Sara Hallin and Aurélien Saghaï jointly finalized the manuscript.</p><p>The authors declare that they have no competing interests.</p><p>The ethics statement is not applicable to this study.</p>","PeriodicalId":100834,"journal":{"name":"Journal of Sustainable Agriculture and Environment","volume":"2 4","pages":"541-545"},"PeriodicalIF":0.0000,"publicationDate":"2023-10-09","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1002/sae2.12073","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"Journal of Sustainable Agriculture and Environment","FirstCategoryId":"1085","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1002/sae2.12073","RegionNum":0,"RegionCategory":null,"ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"","JCRName":"","Score":null,"Total":0}
引用次数: 0
Abstract
Crop production depends on input of nitrogen (N) but because N-use efficiency is low in current conventional cropping systems, farmers fertilize much more than the plants need. More than 50% of added fertilizer N is lost to the environment, mainly as nitrate and gaseous N, that is, dinitrogen, nitrous oxide (N2O), and ammonia (Lassaletta et al., 2014). Apart from deteriorating water quality and negatively impacting biodiversity, a main concern is the emissions of the greenhouse gas N2O. Nitrous oxide exhibits a global warming potential approximately 300 times higher than that of CO2, and the N2O concentration in the atmosphere is increasing at an accelerating rate (Thompson et al., 2019). Anthropogenic sources contribute ca. 45% to global N2O emissions, with direct and indirect emissions from N additions in agriculture accounting for ca. 50% (Tian et al., 2020). The negative consequences of N fertilisation therefore make the global food system a key target to limit climate change (Clark et al., 2020) and allow humanity to remain within a safe operating space of the Earth system.
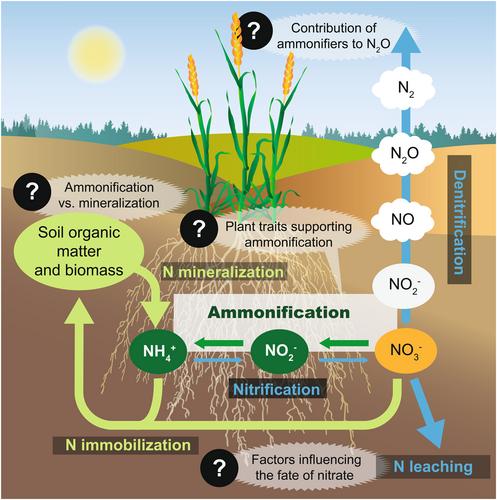
A main challenge for sustainable agriculture is to increase N-use efficiency in cropping systems without compromising yields. One possibility is to improve the retention of soil N by increasing the time N stays in the form of ammonium, as ammonium adsorbs clay particles and soil organic matter. This can be done by using nitrification inhibitors that hinder the microbially mediated oxidation of ammonium to nitrate (Coskun et al., 2017) or by supporting nitrate ammonification, an overlooked process in the N cycle in which nitrate is reduced via nitrite to ammonium (a process also known as dissimilatory nitrate reduction to ammonium [DNRA]). Similar to the competing process of denitrification, nitrate ammonification is performed by phylogenetically diverse microorganisms, which couple the oxidation of various electron donors, most often organic carbon compounds, to the reduction of nitrate under anoxic conditions. Nitrate ammonification creates a short-circuit in the N cycle, bypassing denitrification and N-fixation, and can thereby contribute to primary production (Figure 1). There is, however, a possible risk of ammonia volatilization in alkaline soils. By contrast, the reduction of nitrate to gaseous N oxides through denitrification always results in ecosystem N losses, with a substantial amount emitted as N2O. Thus, the predominant nitrate reduction pathway affects the fate of nitrate and may have major consequences for N-use efficiency in cropping systems and possibly also climate change.
In this commentary, we highlight challenges and key research questions that need to be addressed to be able to evaluate the promises of nitrate ammonification and the feasibility of exploiting this process in sustainable agriculture. These include (i) estimating the relative importance of ammonium production rates via ammonification versus mineralisation, (ii) determining the contribution of nitrate ammonifiers to N2O reduction and production, (iii) assessing the biotic and abiotic factors promoting nitrate ammonification over denitrification, and (iv) exploring the possibility of harnessing plant traits to promote nitrate ammonification and increase N-use efficiency in cropping systems.
Nitrate ammonification rates vary greatly within croplands and managed grasslands (Supporting Information: Table S1). However, it is uncertain how much N it contributes in relation to other sources of ammonium in soil. When comparing these rates to gross N mineralization rates, nitrate ammonification contributes between 0% and 50% (average about 6% and median about 1%; Supporting Information: Table S1) of the produced ammonium in croplands and grasslands, without accounting for the release of previously produced ammonium adsorbed to soil particles. For the comparison, we mainly selected studies based on soil incubation assays with 15N isotopes combined with tracing models using numerical solutions, which allows simultaneous quantification and comparison of multiple N transformations (Rütting et al., 2011). Although these studies suggest that nitrate ammonification could be relevant for sustainable agriculture, the estimates are based on a limited number of studies and soil types as well as different models and assumptions. Further, the concentrations of ammonium and nitrate after substrate addition may also affect the fate of nitrate, considering that nitrate ammonifiers are favoured under low nitrate levels (Saghaï et al., 2023; van den Berg et al., 2016). Additional work is thus needed to gain both a better quantitative understanding of soil N fluxes, also acknowledging sorption and release of ammonium across years in the crop rotation, and a more representative picture of the relative importance of ammonification versus mineralization for delivering ammonium in cropping systems.
One of the possible benefits of enhancing ammonification is the reduction of N2O emissions, as shown by the negative correlation between nitrate ammonification rates and N2O emissions at the ecosystem scale across terrestrial biomes (Cheng et al., 2022). However, only unfertilised soils were included in that analysis, and there is limited information on this relationship in fertilised agricultural soils (Putz et al., 2018). Although the nitrate ammonification pathway in itself should not result in N2O production, small amounts of N2O have been reported from nitrate ammonifier isolates grown under conditions favourable to nitrate ammonification, for example (Stremińska et al., 2012), which has been attributed to detoxification of nitric oxide-mediated by the ammonia forming cytochrome c nitrite reductase, NrfA (Poock et al., 2002). At the same time, several ammonifiers have also been shown to carry nosZ, coding for the N2O reductase, and it has even been discussed that they could play a role as N2O sinks (Hallin et al., 2018). Nevertheless, a recent analysis of ca. 1100 nitrate ammonifier genomes showed that capacity for denitrification and/or nitric oxide reduction is quite common, with 45% carrying one or several denitrification genes and more often for N2O production than reduction (Saghaï et al., 2023). This implies that nitrate ammonifiers play a more complex role in the N cycle than previously thought and underlines that both the community composition of ammonifiers and environmental conditions matter for soil N2O fluxes and the retention of N in agroecosystems.
A recent soil metagenome screening of the gene nrfA, encoding NrfA, revealed that there is a large untapped potential for nitrate ammonification in both croplands and the rhizosphere, only comparable to tropical moist forests (Saghaï et al., 2023). A key issue is how to make the best use of this genetic potential while minimizing the potential for denitrification that typically dominates over that of nitrate ammonification in nearly all terrestrial biomes, especially in croplands (Saghaï et al., 2023). Our understanding of the factors influencing the fate of nitrate in soils is still limited (Cheng et al., 2022), although a few management practices leading to higher soil organic carbon and carbon-to-nitrate ratio, like the inclusion of short-term grasslands in crop rotations, straw incorporation and no-tillage, have been shown to increase nitrate ammonification rates or the relative importance of nitrate ammonification to denitrification at single locales (Putz et al., 2018; Yuan, Liu, et al., 2022; Table 1). However, practices aiming to increase carbon sequestration should also optimize N fertilization strategies, especially in soils with low carbon content (Saghaï et al., 2023). Recent work has highlighted the potential of biochar with high electron shuttle capacity to support nitrate ammonification in rice paddies (Yuan, Wang, et al., 2022), as this pathway requires more electrons to be transferred than denitrification (Tiedje et al., 1982). Although it is more likely to achieve effective N retention in cropping systems if ammonium production and plant uptake are closely coupled, most studies comparing denitrification and ammonification are typically conducted in ‘bulk’ soil and knowledge of plant controls on nitrate ammonification as well as on the competition between the two processes in the root environment is lacking.
Exploiting plant–microbe interactions has been identified as a promising strategy to develop sustainable crops. Large efforts are devoted to identify relevant plant functional traits and their genetic determinants (Bergelson et al., 2021), also for improving N cycling in agroecosystems (Abalos et al., 2019). Plants modify the physical and chemical conditions in the rhizosphere in both time and space, affecting the two major factors regulating the microbial reduction of nitrate, that is, the availability of the main substrates (nitrate and easily accessible organic carbon) and the oxygen partial pressure. Thus, plants with high N uptake efficiency and high growth rates exacerbate the competition for mineral N (Moreau et al., 2015). This could possibly create a more favourable environment for ammonification than for denitrification, as enzymes used in ammonification typically display a lower affinity for nitrate and nitrite (van den Berg et al., 2016). Moreover, the quantity and quality of organic carbon affect not only denitrification rates and the abundance of denitrifying microorganisms (Henry et al., 2008) but also the partitioning of nitrate between the two competing pathways (Carlson et al., 2020), although this has not been demonstrated in planta with natural root exudates. With the rhizosphere being a hotspot for microbial activity, including N assimilation and transformations (Philippot et al., 2013), understanding how plant traits can favour nitrate ammonification over denitrification and allow plants to take advantage of the ammonium produced by ammonifiers will be crucial for the selection and breeding of climate-smart crop varieties (Table 1).
Conservation of soil N as ammonium means transitioning from nitrate to ammonium-based agriculture (Subbarao & Searchinger, 2021). Apart from decreasing N losses, this could also lead to higher crop yields if ammonium-to-nitrate ratios in soil are optimized. Indeed, a mix of the two forms of reactive N can improve crop yields compared with conditions with only nitrate (Wang et al., 2019), likely by making use of the plant's separate systems for the assimilation of ammonium and nitrate and thereby increasing N uptake efficiency. Similarly, ammonium has been shown to decrease plant N uptake if nitrate levels are low (Kuppe & Postma, 2023). Ammonium can also be toxic to plants, although responses to ammonium vary not only among crops but also among crop varieties. This can be exploited for selection and breeding for crops that profit from higher ammonium levels. Improving soil N retention by promoting nitrate ammonification should work hand in hand with other efforts to improve N-use efficiency based on further developing synthetic nitrification inhibitors and biological nitrification inhibition since all these approaches rely on more ammonium-based cropping systems.
Sara Hallin and Aurélien Saghaï conceived the study and acquired funding. Sara Hallin and Aurélien Saghaï compiled the data in Table S1. Sara Hallin wrote the original draft, and Sara Hallin and Aurélien Saghaï jointly finalized the manuscript.
The authors declare that they have no competing interests.
The ethics statement is not applicable to this study.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


