{"title":"立体时空细胞在分子医学中的意义","authors":"Xuanqi Liu, Wanxin Duan, Yuyang Qiu, Ruyi Li, Yuanlin Song, Xiangdong Wang","doi":"10.1002/ctd2.70077","DOIUrl":null,"url":null,"abstract":"<p>With continuous development of biotechnology, our understanding of cells, the fundamental units of the human body and their functions has deepened significantly. The structural and functional characteristics of cells are increasingly recognised as multidimensional and complex, shaped by their tissue and organ locations, intercellular connections, interactions in the extracellular fluids and the dynamics of subcellular organelles and molecules. Cellular heterogeneity among cells primarily arises from variations in the extracellular microenvironment, intracellular genetic diversity, and the spatial and dynamic arrangement of subcellular components, including nuclei, organelles, molecules and cytoplasm. Recent research has evidenced that a large proportion of proteins are spatially allocated in intracellular compartments, including both membrane-bound and membrane-less organelles. This localisation forms protein-driven spatial networks that link these organelles, and orient interconnections among the compartments.<span><sup>1</sup></span> These spatiotemporal distributions of proteins dynamically altered in response to extracellular stimuli and pathogens, regulating protein movements, remodelling and functions independently of mere changes in protein abundance. Trans-compartmental translocations of intracellular components orientate dynamic and multiple regulations of signalling and functions. Studies on the spatiotemporal dynamics of intracellular proteomic and phosphor-proteomic signalling networks have demonstrated that receptor adaptor proteins can be re-distributed among subcellular compartments. This can perform transition from a free cytosolic form to membrane-bound fractions and be targeted to receptors through vesicles, a process activated by the phosphorylation of tyrosine residues in the receptor such as the interaction between epidermal growth factor and epidermal growth factor receptor.<span><sup>2</sup></span> Various subcellular compartmentations are recognised and defined by specific biomarkers at a two-dimensional (2D) level, which partly shows the spatialisation and temporalisation of compartments and molecular relocations in stereological cells (See Figure 1)</p><p>To distinguish this from conventional 1D or 2D spatialisation, we define the concept of the ‘stereologically spatiotemporal cell’ (SST-cell) to describe the precise 3D localisation and interactions of intra- and extracellular components at the single-cell levels. We propose that understanding the SST-cell represents a new frontline in clinical single-cell biomedicine, providing new insights for the clinical translation and application of molecular medicine.<span><sup>3</sup></span> Changes in intracellular components result in a high complexity of intercellular heterogeneity and multidimensional dynamics, posing challenges for real-time monitoring and reproducibility. One of the major challenges is to accurately delineate and interpret the complex multidimensional structures and functions within and surrounding a single cell. Due to the difficulty in detecting the dynamic changes in intracellular components, it remains challenging to uncover and monitor the phenotypic diversity, autonomous responses and metabolic functions of cellular components in their precise stereologically spatiotemporal contexts. A new technology for the genetic tracing codes of distinct endodermal regions was recently developed by integrating high-throughput, high-precision single-cell RNA sequencing with sophisticated imaging.<span><sup>4</sup></span> This approach uncovered the spatiotemporal and genetic lineage differentiation of endodermal cells at single-cell resolution. The spatiotemporal trajectories and dynamic rearrangement of each single cell during early endodermal organogenesis were shown to be influenced by the 3D extracellular microenvironments and intercellular communication. These findings imply that stereological and spatiotemporal regulation may play an important role in organ development via cell recruitment or differentiation from multi-origins. The intra- and extra-SST-cell can be comprehensively presented using a continuous spatial transcriptomic data from tissue sections, such as that obtained by Stereo-seq, combined with computational mapping at single-cell resolution mapping.<span><sup>5</sup></span> The current perspective focuses on the importance of intra- and extracellular stereoscopic changes in SST-cells, the technological advances enabling their precise detection of intracellular positional changes, and their implications for the diagnosis and treatment of human diseases.</p><p>Spatiotemporal changes in 3D chromatin architecture are important for determining transcriptional regulations, genetic epigenetics and the transmission of genetic information within SST-cells. Technological advancements have deepened our understanding of the density and structural changes in 3D chromatin. The dynamic transformation of its spatial structure changes gene expression, which plays an important role in the development and regulation of genetic diseases. The 3D construction of chromosomes ensures their proper formation, and spatial positioning facilitates the recruitment of regulatory factors and underlies the mechanisms by which these factors maintain chromatin architecture. This 3D chromatin architecture can be disrupted by the dysfunction of mediators of replication and DNA double-strand breaks, and impair topologically associating domains rather than DNA synthesis itself, leading to unintended replication events in damaged chromatin and increased DNA damage, particularly in cancer cells.<span><sup>6</sup></span> During cell cycle, the spatiotemporal architecture of chromatin is dynamically regulated by interactions among structural maintenance of chromosome complexes,<span><sup>7</sup></span> including chromatin loop-extruding cohesion, sister chromatid-cohesive cohesion and mitotic chromosome-associated condensins. This 3D chromatin organisation plays a crucial role not only for maintaining transcriptomic network integrity and nuclear shape but also for influencing the spatiotemporal locations of subcellular organelles and the overall structure of the SST-cells. Recently, a new sequencing-based method named linking mRNA to chromatin architecture has enabled the simultaneous measurement of the single-cell 3D genome architecture in the nucleus and transcriptomic profiles in the cytoplasm of the same cell.<span><sup>8</sup></span> This powerful tool in single-cell measurements precisely captures high-order chromatin structure, comprehensively provides the transcriptomic profiles, accurately distinguishes cell types based on chromatin interactions and gene expression, dynamically examines the role of gene positioning in expression, and systematically defines continuous cell-state trajectory during development. Spatiotemporal alterations in 3D chromatin architecture can cause multiple disorders, including defects in organ development, aberrant neural connectivity, carcinogenesis and cancer progression.</p><p>A large number of intracellular organelles and sub-organelles, along with their intercommunications, decides cellular biological types, subtype specification and type-specific functions. The cell molecular phenomes and functions of a cell are fully dependent upon dynamics of organelle numbers, volumes, speeds, positions and dynamic inter-organelle contacts, particularly among membrane-bound organelles. Most organelles and sub-organelles are identified using specific protein markers that define their identity, specificity, spatial separation and abundance in response to microenvironmental or pathological stimuli. The set of all organelle interactions, termed as the organelle interactome, performs molecular exchange and signal transductions, either through direct membrane contacts or proximity-based interactions among organelles. Using two-layer DNA seqFISH+, a method that simultaneously detects genomic loci, transcriptomes and subnuclear structures in the same single cell, Takei et al. demonstrated that repressive chromatin or heterochromatin regions as part of the subnuclear compartments vary in a cell type-dependent pattern.<span><sup>9</sup></span> Within the subnuclear interactome, both RNA polymerase II-enriched and speckle-associated regions showed cell-type-specific gene expression, where the former are locally associated with long, sparsely distributed genes, and the latter with short, densely packed genes. High-resolution, single-cell multi-omic technologies that enable the simultaneous observation of subnuclear structures, corresponding genomic regions, and regulated gene expression in a single-cell within complex tissues provide new insights into the mechanisms of gene dysregulations, misexpressions and dysfunctions in diseases. The characteristic distribution and dispersion pattern of each organelle in 3D space and a reproducible pattern of contacts among organelle interactomes can be changed in response to pathogens, carcinogens and therapies. Among these, various shapes and functions of endoplasmic reticulum (ER) are hardly monitored with dynamical morphology and the cytoplasmic flows in the cell. The adhesion site disassembly, actin dynamics and ER‒plasma membrane polarisation may be regulated by ER‒actin tether-oriented calcium signalling factor and calcium signalling near ER‒actin interfaces.<span><sup>10</sup></span> It is possible that those morphological sub-domains/sub-organelles of ER are prepared for contacting and functioning with other organelles. These mitochondrial and lysosomal dynamics and intercommunications are carrier out and regulated through active, GTP-bound lysosomal RAB7, and can be disconnected by the RAB7 GTPase-activating protein TBC1D15, or fissioned by lysosomal RAB7 hydrolysis via TBC1D15.<span><sup>11</sup></span> Interventions of organelle interactomes cab be a new alternative to discovery of diagnostic biomarkers and therapeutic targets.</p><p>Multiple dimensional dynamics of intra- and extracellular components decide the cell performances in the tissue and organ functions in the body. The development of technologies such as continuous Stereo-seq, a combination of DNA nanoball-patterned arrays and in situ RNA capture, allows the 3D locations and contacts of intra- and extracellular components to be defined with high-spatial-resolution omics sequencing. This spatial omics-sequencing approach enables the detailed dissection of spatial heterogeneity among different cell subtypes and states, as well as their intercellular communications.<span><sup>5</sup></span> Using continuous spatial transcriptomics, their 3D lineage trajectories of intra- and extra-embryonic molecular and cellular components have been traced, revealing how their interactions contribute to early development.<span><sup>12</sup></span> The development of the human embryo critically depends upon characters and patterns of 3D spatial arrangements of those cells, while the intracellular transcriptomic regulations orient their positioning and function of these cells. By integrating experimental and computational tools for the efficient and special capture of continuous spatial transcriptomes, the metastatic capacity of a tumour has been found to be highly dependent on the spatial rearrangements and contacts of various cell types within the tumour microenvironment.<span><sup>13</sup></span> This spatially resolved approach provides a potential to determine the stereological spatialisation of intra- and extracellular components, map 3D cell‒cell interactions with simultaneous states of transcriptomic regulatory networks, and explore the molecular mechanism based on cellular spatial re-locations. A spatial atlas of the human thymus, for example, has demonstrated the full 3D trajectory of T-cell lineage differentiation at beginning of the second trimester of foetal development, including cytokine and chemokine expression patterns, the spatial shift of thymic epithelial cell populations and subtypes between medullary and cortical regions, and development of CD4 and CD8 thymocytes.<span><sup>14</sup></span> In tumours, the stereological spatialisation of tumour microenvironment provides 3D locations, contact interface and communication networks among cancer cells, cancer-associated cells, immune cells and signalling mediators. These insights are obtained by measuring the correlations between average module expression, sample purity and spatial metrics such as the relative distance between centroid and periphery.<span><sup>15</sup></span> In contrast to 1D and 2D spatial transcriptomics, 3D spatial profiling of the tumour microenvironment exhibits more specific and precise interactions among activated cells, populations trajectories, cell subtypes and spatial proclivities of interactomes and signalling pathways. Moreover, the volume of tumour microenvironment is spatially divided into connectivity, loop and microregion of tumour growth patterns and interactomes. Spatial distributions of 3D tumour volume decide the nature of cancer cell identity and metastasis.<span><sup>16</sup></span></p><p>The SST-cell is a new and important frontline of molecular medicine, an unexplored resource for uncovering diagnostic biomarkers and therapeutic targets, and an emerging discipline of biomedicine. A major challenge is to precisely define and dynamically monitor the stereological spatialisation and its changes in intra- and extracellular components. Although organelle-specific marker proteins can help evaluate the interactions between them, it remains technically difficult to accurately define the 3D locations and distances between organelles, especially at the simultaneous time point these interactions occur. There is an urgent need to develop sub-organelle-specific antibody panels for distinguishing between sub-organellar compartments of the organelle and map their multidirectional spatial orientations within the cell. In addition to the cost burden of continuous Stereo-seq, standardised analyses and graphical representations in 3D are critical to overcome the overlap of data points and patterns due to the high-throughput manners of spatial multi-omics. New, precise and repeatable methodologies should be established to address the multilayered spatial organisation and functions of intracellular and intraorganellar molecular components. Further studies on human cellular and organellar interactomes are expected, especially in pathological contexts. Resolving the 3D spatial locations and functional states of intracellular components at single-cell resolution remains a particularly formidable challenge.</p><p>The clinical translation of SST-cell measurements requires a clear cell identity annotation, simplified experimental workflows, artificial intelligence-driven high-throughput analysis, uniform sampling, adaptability to diverse cell sizes and broad technical extensibility. To this end, a spatially enhanced resolution single-cell sequencing platform, Stereo-cell, is developed. Based on high-density DNA nanoball-patterned arrays, it is able to measure cell surface proteins, morphologies and transcriptomic profiles simultaneously.<span><sup>17</sup></span> In addition to using marker gene panels for annotation, this platform allows scattered cells seeded on poly-L-lysine-coated array chips to be stained and labelled with biochemicals or target-specific antibodies, similar to routine applications in clinical haematology. This system integrates imaging-based measurements with molecular multi-omic profiles, and can accommodate extracellular vesicles, microstructures, large multinucleated cells and other complex clinical specimens for in situ multi-omic profiling.</p><p>A deep understanding of cellular and organellar interactomes and their real-time communication requires monitoring the stereological spatialisations of intracellular components in live cells. A system of CRISPR-mediated transcriptome organisation (CRISPR-TO) was developed for real-time monitoring of the spatial dynamics of RNA location across various subcellular compartments in living cells.<span><sup>18</sup></span> CRISPR-TO couples the dCas13 unit (fused with one ABA dimerisation domain) with the signal unit (subcellular localisation signal or motor protein fused with the other dimerisation domain) through the gRNAs unit (chemical-inducible dimerisation) to perform the programmable control of endogenous RNA localisation in live cells. CRISPR-TO is applicable in primary human cells, supports multiplex detection of RNA localisations to investigate cooperative roles, and is capable of programmability to screen high-throughput functions. In addition to RNA levels, a ‘zero-distance’ photo-crosslinking approach has been developed to identify the proteome that physically interacts with DNA in living cells.<span><sup>19</sup></span> This creates new opportunities to define the manner of protein‒DNA interactions, maintenance of 3D chromatin architecture, and link the stereological spatialisation of genomic regulation, and relations with cellular and molecular phenomes and functions.</p><p>In conclusion, cell function and morphology are highly dependent upon the stereologically spatiotemporal positioning, precise dimensions and interactions of subcellular organelles. The stability of the 3D chromatin architecture, along with spatial intra- and extracellular and organellar interactomes, governs cell differentiation and maintains the bioecology of microenvironments. The multi-omic profiles and locations in a SST-cell can be defined by combining image- and molecular omics-based strategies and by monitoring the real-time dynamics of spatial transcriptomes and proteomics, although technical hurdles remain. SST-cell biology provides a transformative new perspective for understanding pathogenesis and opens new alternatives to discovery of biomarkers for diagnosis and drug development.</p><p>XQL and WXD contribute to the data collections and manuscript writing; YYQ and RYL are responsible for the literature and discussion; YLS and XDW design the therory, provide the systemic idea as well as the manuscript writing.</p><p>None.</p>","PeriodicalId":72605,"journal":{"name":"Clinical and translational discovery","volume":"5 4","pages":""},"PeriodicalIF":1.9000,"publicationDate":"2025-07-27","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1002/ctd2.70077","citationCount":"0","resultStr":"{\"title\":\"Significance of stereologically spatiotemporal cells in molecular medicine\",\"authors\":\"Xuanqi Liu, Wanxin Duan, Yuyang Qiu, Ruyi Li, Yuanlin Song, Xiangdong Wang\",\"doi\":\"10.1002/ctd2.70077\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p>With continuous development of biotechnology, our understanding of cells, the fundamental units of the human body and their functions has deepened significantly. The structural and functional characteristics of cells are increasingly recognised as multidimensional and complex, shaped by their tissue and organ locations, intercellular connections, interactions in the extracellular fluids and the dynamics of subcellular organelles and molecules. Cellular heterogeneity among cells primarily arises from variations in the extracellular microenvironment, intracellular genetic diversity, and the spatial and dynamic arrangement of subcellular components, including nuclei, organelles, molecules and cytoplasm. Recent research has evidenced that a large proportion of proteins are spatially allocated in intracellular compartments, including both membrane-bound and membrane-less organelles. This localisation forms protein-driven spatial networks that link these organelles, and orient interconnections among the compartments.<span><sup>1</sup></span> These spatiotemporal distributions of proteins dynamically altered in response to extracellular stimuli and pathogens, regulating protein movements, remodelling and functions independently of mere changes in protein abundance. Trans-compartmental translocations of intracellular components orientate dynamic and multiple regulations of signalling and functions. Studies on the spatiotemporal dynamics of intracellular proteomic and phosphor-proteomic signalling networks have demonstrated that receptor adaptor proteins can be re-distributed among subcellular compartments. This can perform transition from a free cytosolic form to membrane-bound fractions and be targeted to receptors through vesicles, a process activated by the phosphorylation of tyrosine residues in the receptor such as the interaction between epidermal growth factor and epidermal growth factor receptor.<span><sup>2</sup></span> Various subcellular compartmentations are recognised and defined by specific biomarkers at a two-dimensional (2D) level, which partly shows the spatialisation and temporalisation of compartments and molecular relocations in stereological cells (See Figure 1)</p><p>To distinguish this from conventional 1D or 2D spatialisation, we define the concept of the ‘stereologically spatiotemporal cell’ (SST-cell) to describe the precise 3D localisation and interactions of intra- and extracellular components at the single-cell levels. We propose that understanding the SST-cell represents a new frontline in clinical single-cell biomedicine, providing new insights for the clinical translation and application of molecular medicine.<span><sup>3</sup></span> Changes in intracellular components result in a high complexity of intercellular heterogeneity and multidimensional dynamics, posing challenges for real-time monitoring and reproducibility. One of the major challenges is to accurately delineate and interpret the complex multidimensional structures and functions within and surrounding a single cell. Due to the difficulty in detecting the dynamic changes in intracellular components, it remains challenging to uncover and monitor the phenotypic diversity, autonomous responses and metabolic functions of cellular components in their precise stereologically spatiotemporal contexts. A new technology for the genetic tracing codes of distinct endodermal regions was recently developed by integrating high-throughput, high-precision single-cell RNA sequencing with sophisticated imaging.<span><sup>4</sup></span> This approach uncovered the spatiotemporal and genetic lineage differentiation of endodermal cells at single-cell resolution. The spatiotemporal trajectories and dynamic rearrangement of each single cell during early endodermal organogenesis were shown to be influenced by the 3D extracellular microenvironments and intercellular communication. These findings imply that stereological and spatiotemporal regulation may play an important role in organ development via cell recruitment or differentiation from multi-origins. The intra- and extra-SST-cell can be comprehensively presented using a continuous spatial transcriptomic data from tissue sections, such as that obtained by Stereo-seq, combined with computational mapping at single-cell resolution mapping.<span><sup>5</sup></span> The current perspective focuses on the importance of intra- and extracellular stereoscopic changes in SST-cells, the technological advances enabling their precise detection of intracellular positional changes, and their implications for the diagnosis and treatment of human diseases.</p><p>Spatiotemporal changes in 3D chromatin architecture are important for determining transcriptional regulations, genetic epigenetics and the transmission of genetic information within SST-cells. Technological advancements have deepened our understanding of the density and structural changes in 3D chromatin. The dynamic transformation of its spatial structure changes gene expression, which plays an important role in the development and regulation of genetic diseases. The 3D construction of chromosomes ensures their proper formation, and spatial positioning facilitates the recruitment of regulatory factors and underlies the mechanisms by which these factors maintain chromatin architecture. This 3D chromatin architecture can be disrupted by the dysfunction of mediators of replication and DNA double-strand breaks, and impair topologically associating domains rather than DNA synthesis itself, leading to unintended replication events in damaged chromatin and increased DNA damage, particularly in cancer cells.<span><sup>6</sup></span> During cell cycle, the spatiotemporal architecture of chromatin is dynamically regulated by interactions among structural maintenance of chromosome complexes,<span><sup>7</sup></span> including chromatin loop-extruding cohesion, sister chromatid-cohesive cohesion and mitotic chromosome-associated condensins. This 3D chromatin organisation plays a crucial role not only for maintaining transcriptomic network integrity and nuclear shape but also for influencing the spatiotemporal locations of subcellular organelles and the overall structure of the SST-cells. Recently, a new sequencing-based method named linking mRNA to chromatin architecture has enabled the simultaneous measurement of the single-cell 3D genome architecture in the nucleus and transcriptomic profiles in the cytoplasm of the same cell.<span><sup>8</sup></span> This powerful tool in single-cell measurements precisely captures high-order chromatin structure, comprehensively provides the transcriptomic profiles, accurately distinguishes cell types based on chromatin interactions and gene expression, dynamically examines the role of gene positioning in expression, and systematically defines continuous cell-state trajectory during development. Spatiotemporal alterations in 3D chromatin architecture can cause multiple disorders, including defects in organ development, aberrant neural connectivity, carcinogenesis and cancer progression.</p><p>A large number of intracellular organelles and sub-organelles, along with their intercommunications, decides cellular biological types, subtype specification and type-specific functions. The cell molecular phenomes and functions of a cell are fully dependent upon dynamics of organelle numbers, volumes, speeds, positions and dynamic inter-organelle contacts, particularly among membrane-bound organelles. Most organelles and sub-organelles are identified using specific protein markers that define their identity, specificity, spatial separation and abundance in response to microenvironmental or pathological stimuli. The set of all organelle interactions, termed as the organelle interactome, performs molecular exchange and signal transductions, either through direct membrane contacts or proximity-based interactions among organelles. Using two-layer DNA seqFISH+, a method that simultaneously detects genomic loci, transcriptomes and subnuclear structures in the same single cell, Takei et al. demonstrated that repressive chromatin or heterochromatin regions as part of the subnuclear compartments vary in a cell type-dependent pattern.<span><sup>9</sup></span> Within the subnuclear interactome, both RNA polymerase II-enriched and speckle-associated regions showed cell-type-specific gene expression, where the former are locally associated with long, sparsely distributed genes, and the latter with short, densely packed genes. High-resolution, single-cell multi-omic technologies that enable the simultaneous observation of subnuclear structures, corresponding genomic regions, and regulated gene expression in a single-cell within complex tissues provide new insights into the mechanisms of gene dysregulations, misexpressions and dysfunctions in diseases. The characteristic distribution and dispersion pattern of each organelle in 3D space and a reproducible pattern of contacts among organelle interactomes can be changed in response to pathogens, carcinogens and therapies. Among these, various shapes and functions of endoplasmic reticulum (ER) are hardly monitored with dynamical morphology and the cytoplasmic flows in the cell. The adhesion site disassembly, actin dynamics and ER‒plasma membrane polarisation may be regulated by ER‒actin tether-oriented calcium signalling factor and calcium signalling near ER‒actin interfaces.<span><sup>10</sup></span> It is possible that those morphological sub-domains/sub-organelles of ER are prepared for contacting and functioning with other organelles. These mitochondrial and lysosomal dynamics and intercommunications are carrier out and regulated through active, GTP-bound lysosomal RAB7, and can be disconnected by the RAB7 GTPase-activating protein TBC1D15, or fissioned by lysosomal RAB7 hydrolysis via TBC1D15.<span><sup>11</sup></span> Interventions of organelle interactomes cab be a new alternative to discovery of diagnostic biomarkers and therapeutic targets.</p><p>Multiple dimensional dynamics of intra- and extracellular components decide the cell performances in the tissue and organ functions in the body. The development of technologies such as continuous Stereo-seq, a combination of DNA nanoball-patterned arrays and in situ RNA capture, allows the 3D locations and contacts of intra- and extracellular components to be defined with high-spatial-resolution omics sequencing. This spatial omics-sequencing approach enables the detailed dissection of spatial heterogeneity among different cell subtypes and states, as well as their intercellular communications.<span><sup>5</sup></span> Using continuous spatial transcriptomics, their 3D lineage trajectories of intra- and extra-embryonic molecular and cellular components have been traced, revealing how their interactions contribute to early development.<span><sup>12</sup></span> The development of the human embryo critically depends upon characters and patterns of 3D spatial arrangements of those cells, while the intracellular transcriptomic regulations orient their positioning and function of these cells. By integrating experimental and computational tools for the efficient and special capture of continuous spatial transcriptomes, the metastatic capacity of a tumour has been found to be highly dependent on the spatial rearrangements and contacts of various cell types within the tumour microenvironment.<span><sup>13</sup></span> This spatially resolved approach provides a potential to determine the stereological spatialisation of intra- and extracellular components, map 3D cell‒cell interactions with simultaneous states of transcriptomic regulatory networks, and explore the molecular mechanism based on cellular spatial re-locations. A spatial atlas of the human thymus, for example, has demonstrated the full 3D trajectory of T-cell lineage differentiation at beginning of the second trimester of foetal development, including cytokine and chemokine expression patterns, the spatial shift of thymic epithelial cell populations and subtypes between medullary and cortical regions, and development of CD4 and CD8 thymocytes.<span><sup>14</sup></span> In tumours, the stereological spatialisation of tumour microenvironment provides 3D locations, contact interface and communication networks among cancer cells, cancer-associated cells, immune cells and signalling mediators. These insights are obtained by measuring the correlations between average module expression, sample purity and spatial metrics such as the relative distance between centroid and periphery.<span><sup>15</sup></span> In contrast to 1D and 2D spatial transcriptomics, 3D spatial profiling of the tumour microenvironment exhibits more specific and precise interactions among activated cells, populations trajectories, cell subtypes and spatial proclivities of interactomes and signalling pathways. Moreover, the volume of tumour microenvironment is spatially divided into connectivity, loop and microregion of tumour growth patterns and interactomes. Spatial distributions of 3D tumour volume decide the nature of cancer cell identity and metastasis.<span><sup>16</sup></span></p><p>The SST-cell is a new and important frontline of molecular medicine, an unexplored resource for uncovering diagnostic biomarkers and therapeutic targets, and an emerging discipline of biomedicine. A major challenge is to precisely define and dynamically monitor the stereological spatialisation and its changes in intra- and extracellular components. Although organelle-specific marker proteins can help evaluate the interactions between them, it remains technically difficult to accurately define the 3D locations and distances between organelles, especially at the simultaneous time point these interactions occur. There is an urgent need to develop sub-organelle-specific antibody panels for distinguishing between sub-organellar compartments of the organelle and map their multidirectional spatial orientations within the cell. In addition to the cost burden of continuous Stereo-seq, standardised analyses and graphical representations in 3D are critical to overcome the overlap of data points and patterns due to the high-throughput manners of spatial multi-omics. New, precise and repeatable methodologies should be established to address the multilayered spatial organisation and functions of intracellular and intraorganellar molecular components. Further studies on human cellular and organellar interactomes are expected, especially in pathological contexts. Resolving the 3D spatial locations and functional states of intracellular components at single-cell resolution remains a particularly formidable challenge.</p><p>The clinical translation of SST-cell measurements requires a clear cell identity annotation, simplified experimental workflows, artificial intelligence-driven high-throughput analysis, uniform sampling, adaptability to diverse cell sizes and broad technical extensibility. To this end, a spatially enhanced resolution single-cell sequencing platform, Stereo-cell, is developed. Based on high-density DNA nanoball-patterned arrays, it is able to measure cell surface proteins, morphologies and transcriptomic profiles simultaneously.<span><sup>17</sup></span> In addition to using marker gene panels for annotation, this platform allows scattered cells seeded on poly-L-lysine-coated array chips to be stained and labelled with biochemicals or target-specific antibodies, similar to routine applications in clinical haematology. This system integrates imaging-based measurements with molecular multi-omic profiles, and can accommodate extracellular vesicles, microstructures, large multinucleated cells and other complex clinical specimens for in situ multi-omic profiling.</p><p>A deep understanding of cellular and organellar interactomes and their real-time communication requires monitoring the stereological spatialisations of intracellular components in live cells. A system of CRISPR-mediated transcriptome organisation (CRISPR-TO) was developed for real-time monitoring of the spatial dynamics of RNA location across various subcellular compartments in living cells.<span><sup>18</sup></span> CRISPR-TO couples the dCas13 unit (fused with one ABA dimerisation domain) with the signal unit (subcellular localisation signal or motor protein fused with the other dimerisation domain) through the gRNAs unit (chemical-inducible dimerisation) to perform the programmable control of endogenous RNA localisation in live cells. CRISPR-TO is applicable in primary human cells, supports multiplex detection of RNA localisations to investigate cooperative roles, and is capable of programmability to screen high-throughput functions. In addition to RNA levels, a ‘zero-distance’ photo-crosslinking approach has been developed to identify the proteome that physically interacts with DNA in living cells.<span><sup>19</sup></span> This creates new opportunities to define the manner of protein‒DNA interactions, maintenance of 3D chromatin architecture, and link the stereological spatialisation of genomic regulation, and relations with cellular and molecular phenomes and functions.</p><p>In conclusion, cell function and morphology are highly dependent upon the stereologically spatiotemporal positioning, precise dimensions and interactions of subcellular organelles. The stability of the 3D chromatin architecture, along with spatial intra- and extracellular and organellar interactomes, governs cell differentiation and maintains the bioecology of microenvironments. The multi-omic profiles and locations in a SST-cell can be defined by combining image- and molecular omics-based strategies and by monitoring the real-time dynamics of spatial transcriptomes and proteomics, although technical hurdles remain. SST-cell biology provides a transformative new perspective for understanding pathogenesis and opens new alternatives to discovery of biomarkers for diagnosis and drug development.</p><p>XQL and WXD contribute to the data collections and manuscript writing; YYQ and RYL are responsible for the literature and discussion; YLS and XDW design the therory, provide the systemic idea as well as the manuscript writing.</p><p>None.</p>\",\"PeriodicalId\":72605,\"journal\":{\"name\":\"Clinical and translational discovery\",\"volume\":\"5 4\",\"pages\":\"\"},\"PeriodicalIF\":1.9000,\"publicationDate\":\"2025-07-27\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://onlinelibrary.wiley.com/doi/epdf/10.1002/ctd2.70077\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"Clinical and translational discovery\",\"FirstCategoryId\":\"1085\",\"ListUrlMain\":\"https://onlinelibrary.wiley.com/doi/10.1002/ctd2.70077\",\"RegionNum\":0,\"RegionCategory\":null,\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"\",\"JCRName\":\"\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"Clinical and translational discovery","FirstCategoryId":"1085","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1002/ctd2.70077","RegionNum":0,"RegionCategory":null,"ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"","JCRName":"","Score":null,"Total":0}
Significance of stereologically spatiotemporal cells in molecular medicine
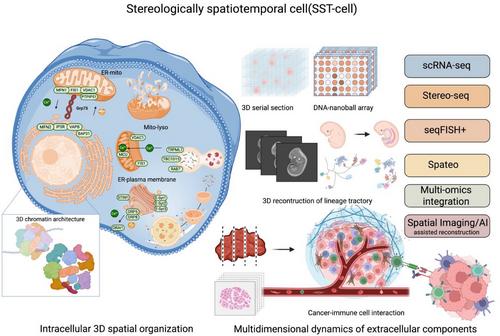
With continuous development of biotechnology, our understanding of cells, the fundamental units of the human body and their functions has deepened significantly. The structural and functional characteristics of cells are increasingly recognised as multidimensional and complex, shaped by their tissue and organ locations, intercellular connections, interactions in the extracellular fluids and the dynamics of subcellular organelles and molecules. Cellular heterogeneity among cells primarily arises from variations in the extracellular microenvironment, intracellular genetic diversity, and the spatial and dynamic arrangement of subcellular components, including nuclei, organelles, molecules and cytoplasm. Recent research has evidenced that a large proportion of proteins are spatially allocated in intracellular compartments, including both membrane-bound and membrane-less organelles. This localisation forms protein-driven spatial networks that link these organelles, and orient interconnections among the compartments.1 These spatiotemporal distributions of proteins dynamically altered in response to extracellular stimuli and pathogens, regulating protein movements, remodelling and functions independently of mere changes in protein abundance. Trans-compartmental translocations of intracellular components orientate dynamic and multiple regulations of signalling and functions. Studies on the spatiotemporal dynamics of intracellular proteomic and phosphor-proteomic signalling networks have demonstrated that receptor adaptor proteins can be re-distributed among subcellular compartments. This can perform transition from a free cytosolic form to membrane-bound fractions and be targeted to receptors through vesicles, a process activated by the phosphorylation of tyrosine residues in the receptor such as the interaction between epidermal growth factor and epidermal growth factor receptor.2 Various subcellular compartmentations are recognised and defined by specific biomarkers at a two-dimensional (2D) level, which partly shows the spatialisation and temporalisation of compartments and molecular relocations in stereological cells (See Figure 1)
To distinguish this from conventional 1D or 2D spatialisation, we define the concept of the ‘stereologically spatiotemporal cell’ (SST-cell) to describe the precise 3D localisation and interactions of intra- and extracellular components at the single-cell levels. We propose that understanding the SST-cell represents a new frontline in clinical single-cell biomedicine, providing new insights for the clinical translation and application of molecular medicine.3 Changes in intracellular components result in a high complexity of intercellular heterogeneity and multidimensional dynamics, posing challenges for real-time monitoring and reproducibility. One of the major challenges is to accurately delineate and interpret the complex multidimensional structures and functions within and surrounding a single cell. Due to the difficulty in detecting the dynamic changes in intracellular components, it remains challenging to uncover and monitor the phenotypic diversity, autonomous responses and metabolic functions of cellular components in their precise stereologically spatiotemporal contexts. A new technology for the genetic tracing codes of distinct endodermal regions was recently developed by integrating high-throughput, high-precision single-cell RNA sequencing with sophisticated imaging.4 This approach uncovered the spatiotemporal and genetic lineage differentiation of endodermal cells at single-cell resolution. The spatiotemporal trajectories and dynamic rearrangement of each single cell during early endodermal organogenesis were shown to be influenced by the 3D extracellular microenvironments and intercellular communication. These findings imply that stereological and spatiotemporal regulation may play an important role in organ development via cell recruitment or differentiation from multi-origins. The intra- and extra-SST-cell can be comprehensively presented using a continuous spatial transcriptomic data from tissue sections, such as that obtained by Stereo-seq, combined with computational mapping at single-cell resolution mapping.5 The current perspective focuses on the importance of intra- and extracellular stereoscopic changes in SST-cells, the technological advances enabling their precise detection of intracellular positional changes, and their implications for the diagnosis and treatment of human diseases.
Spatiotemporal changes in 3D chromatin architecture are important for determining transcriptional regulations, genetic epigenetics and the transmission of genetic information within SST-cells. Technological advancements have deepened our understanding of the density and structural changes in 3D chromatin. The dynamic transformation of its spatial structure changes gene expression, which plays an important role in the development and regulation of genetic diseases. The 3D construction of chromosomes ensures their proper formation, and spatial positioning facilitates the recruitment of regulatory factors and underlies the mechanisms by which these factors maintain chromatin architecture. This 3D chromatin architecture can be disrupted by the dysfunction of mediators of replication and DNA double-strand breaks, and impair topologically associating domains rather than DNA synthesis itself, leading to unintended replication events in damaged chromatin and increased DNA damage, particularly in cancer cells.6 During cell cycle, the spatiotemporal architecture of chromatin is dynamically regulated by interactions among structural maintenance of chromosome complexes,7 including chromatin loop-extruding cohesion, sister chromatid-cohesive cohesion and mitotic chromosome-associated condensins. This 3D chromatin organisation plays a crucial role not only for maintaining transcriptomic network integrity and nuclear shape but also for influencing the spatiotemporal locations of subcellular organelles and the overall structure of the SST-cells. Recently, a new sequencing-based method named linking mRNA to chromatin architecture has enabled the simultaneous measurement of the single-cell 3D genome architecture in the nucleus and transcriptomic profiles in the cytoplasm of the same cell.8 This powerful tool in single-cell measurements precisely captures high-order chromatin structure, comprehensively provides the transcriptomic profiles, accurately distinguishes cell types based on chromatin interactions and gene expression, dynamically examines the role of gene positioning in expression, and systematically defines continuous cell-state trajectory during development. Spatiotemporal alterations in 3D chromatin architecture can cause multiple disorders, including defects in organ development, aberrant neural connectivity, carcinogenesis and cancer progression.
A large number of intracellular organelles and sub-organelles, along with their intercommunications, decides cellular biological types, subtype specification and type-specific functions. The cell molecular phenomes and functions of a cell are fully dependent upon dynamics of organelle numbers, volumes, speeds, positions and dynamic inter-organelle contacts, particularly among membrane-bound organelles. Most organelles and sub-organelles are identified using specific protein markers that define their identity, specificity, spatial separation and abundance in response to microenvironmental or pathological stimuli. The set of all organelle interactions, termed as the organelle interactome, performs molecular exchange and signal transductions, either through direct membrane contacts or proximity-based interactions among organelles. Using two-layer DNA seqFISH+, a method that simultaneously detects genomic loci, transcriptomes and subnuclear structures in the same single cell, Takei et al. demonstrated that repressive chromatin or heterochromatin regions as part of the subnuclear compartments vary in a cell type-dependent pattern.9 Within the subnuclear interactome, both RNA polymerase II-enriched and speckle-associated regions showed cell-type-specific gene expression, where the former are locally associated with long, sparsely distributed genes, and the latter with short, densely packed genes. High-resolution, single-cell multi-omic technologies that enable the simultaneous observation of subnuclear structures, corresponding genomic regions, and regulated gene expression in a single-cell within complex tissues provide new insights into the mechanisms of gene dysregulations, misexpressions and dysfunctions in diseases. The characteristic distribution and dispersion pattern of each organelle in 3D space and a reproducible pattern of contacts among organelle interactomes can be changed in response to pathogens, carcinogens and therapies. Among these, various shapes and functions of endoplasmic reticulum (ER) are hardly monitored with dynamical morphology and the cytoplasmic flows in the cell. The adhesion site disassembly, actin dynamics and ER‒plasma membrane polarisation may be regulated by ER‒actin tether-oriented calcium signalling factor and calcium signalling near ER‒actin interfaces.10 It is possible that those morphological sub-domains/sub-organelles of ER are prepared for contacting and functioning with other organelles. These mitochondrial and lysosomal dynamics and intercommunications are carrier out and regulated through active, GTP-bound lysosomal RAB7, and can be disconnected by the RAB7 GTPase-activating protein TBC1D15, or fissioned by lysosomal RAB7 hydrolysis via TBC1D15.11 Interventions of organelle interactomes cab be a new alternative to discovery of diagnostic biomarkers and therapeutic targets.
Multiple dimensional dynamics of intra- and extracellular components decide the cell performances in the tissue and organ functions in the body. The development of technologies such as continuous Stereo-seq, a combination of DNA nanoball-patterned arrays and in situ RNA capture, allows the 3D locations and contacts of intra- and extracellular components to be defined with high-spatial-resolution omics sequencing. This spatial omics-sequencing approach enables the detailed dissection of spatial heterogeneity among different cell subtypes and states, as well as their intercellular communications.5 Using continuous spatial transcriptomics, their 3D lineage trajectories of intra- and extra-embryonic molecular and cellular components have been traced, revealing how their interactions contribute to early development.12 The development of the human embryo critically depends upon characters and patterns of 3D spatial arrangements of those cells, while the intracellular transcriptomic regulations orient their positioning and function of these cells. By integrating experimental and computational tools for the efficient and special capture of continuous spatial transcriptomes, the metastatic capacity of a tumour has been found to be highly dependent on the spatial rearrangements and contacts of various cell types within the tumour microenvironment.13 This spatially resolved approach provides a potential to determine the stereological spatialisation of intra- and extracellular components, map 3D cell‒cell interactions with simultaneous states of transcriptomic regulatory networks, and explore the molecular mechanism based on cellular spatial re-locations. A spatial atlas of the human thymus, for example, has demonstrated the full 3D trajectory of T-cell lineage differentiation at beginning of the second trimester of foetal development, including cytokine and chemokine expression patterns, the spatial shift of thymic epithelial cell populations and subtypes between medullary and cortical regions, and development of CD4 and CD8 thymocytes.14 In tumours, the stereological spatialisation of tumour microenvironment provides 3D locations, contact interface and communication networks among cancer cells, cancer-associated cells, immune cells and signalling mediators. These insights are obtained by measuring the correlations between average module expression, sample purity and spatial metrics such as the relative distance between centroid and periphery.15 In contrast to 1D and 2D spatial transcriptomics, 3D spatial profiling of the tumour microenvironment exhibits more specific and precise interactions among activated cells, populations trajectories, cell subtypes and spatial proclivities of interactomes and signalling pathways. Moreover, the volume of tumour microenvironment is spatially divided into connectivity, loop and microregion of tumour growth patterns and interactomes. Spatial distributions of 3D tumour volume decide the nature of cancer cell identity and metastasis.16
The SST-cell is a new and important frontline of molecular medicine, an unexplored resource for uncovering diagnostic biomarkers and therapeutic targets, and an emerging discipline of biomedicine. A major challenge is to precisely define and dynamically monitor the stereological spatialisation and its changes in intra- and extracellular components. Although organelle-specific marker proteins can help evaluate the interactions between them, it remains technically difficult to accurately define the 3D locations and distances between organelles, especially at the simultaneous time point these interactions occur. There is an urgent need to develop sub-organelle-specific antibody panels for distinguishing between sub-organellar compartments of the organelle and map their multidirectional spatial orientations within the cell. In addition to the cost burden of continuous Stereo-seq, standardised analyses and graphical representations in 3D are critical to overcome the overlap of data points and patterns due to the high-throughput manners of spatial multi-omics. New, precise and repeatable methodologies should be established to address the multilayered spatial organisation and functions of intracellular and intraorganellar molecular components. Further studies on human cellular and organellar interactomes are expected, especially in pathological contexts. Resolving the 3D spatial locations and functional states of intracellular components at single-cell resolution remains a particularly formidable challenge.
The clinical translation of SST-cell measurements requires a clear cell identity annotation, simplified experimental workflows, artificial intelligence-driven high-throughput analysis, uniform sampling, adaptability to diverse cell sizes and broad technical extensibility. To this end, a spatially enhanced resolution single-cell sequencing platform, Stereo-cell, is developed. Based on high-density DNA nanoball-patterned arrays, it is able to measure cell surface proteins, morphologies and transcriptomic profiles simultaneously.17 In addition to using marker gene panels for annotation, this platform allows scattered cells seeded on poly-L-lysine-coated array chips to be stained and labelled with biochemicals or target-specific antibodies, similar to routine applications in clinical haematology. This system integrates imaging-based measurements with molecular multi-omic profiles, and can accommodate extracellular vesicles, microstructures, large multinucleated cells and other complex clinical specimens for in situ multi-omic profiling.
A deep understanding of cellular and organellar interactomes and their real-time communication requires monitoring the stereological spatialisations of intracellular components in live cells. A system of CRISPR-mediated transcriptome organisation (CRISPR-TO) was developed for real-time monitoring of the spatial dynamics of RNA location across various subcellular compartments in living cells.18 CRISPR-TO couples the dCas13 unit (fused with one ABA dimerisation domain) with the signal unit (subcellular localisation signal or motor protein fused with the other dimerisation domain) through the gRNAs unit (chemical-inducible dimerisation) to perform the programmable control of endogenous RNA localisation in live cells. CRISPR-TO is applicable in primary human cells, supports multiplex detection of RNA localisations to investigate cooperative roles, and is capable of programmability to screen high-throughput functions. In addition to RNA levels, a ‘zero-distance’ photo-crosslinking approach has been developed to identify the proteome that physically interacts with DNA in living cells.19 This creates new opportunities to define the manner of protein‒DNA interactions, maintenance of 3D chromatin architecture, and link the stereological spatialisation of genomic regulation, and relations with cellular and molecular phenomes and functions.
In conclusion, cell function and morphology are highly dependent upon the stereologically spatiotemporal positioning, precise dimensions and interactions of subcellular organelles. The stability of the 3D chromatin architecture, along with spatial intra- and extracellular and organellar interactomes, governs cell differentiation and maintains the bioecology of microenvironments. The multi-omic profiles and locations in a SST-cell can be defined by combining image- and molecular omics-based strategies and by monitoring the real-time dynamics of spatial transcriptomes and proteomics, although technical hurdles remain. SST-cell biology provides a transformative new perspective for understanding pathogenesis and opens new alternatives to discovery of biomarkers for diagnosis and drug development.
XQL and WXD contribute to the data collections and manuscript writing; YYQ and RYL are responsible for the literature and discussion; YLS and XDW design the therory, provide the systemic idea as well as the manuscript writing.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


