{"title":"H5N1 禽流感的现状及对人类的风险。","authors":"Michelle Wille, Ian G. Barr","doi":"10.1111/imj.16550","DOIUrl":null,"url":null,"abstract":"<p>Influenza in humans is mainly caused by two different viruses: influenza A virus (IAV) and influenza B virus. Beyond humans, IAV infects a broader range of species, including avian, equine, porcine and canine species. In contrast, influenza B virus almost exclusively infects humans. Avian IAVs are categorised in two main ways: by subtype and by pathotype. Subtyping is based on the diversity of the two major surface proteins: haemagglutinin (HA) and neuraminidase (NA). These proteins are highly diverse, with 19 HA subtypes and 11 NA subtypes, where the HA-NA combination added together make up the basic identity tag, or subtype, for example, H1N1 and H5N1. To date, only three IAV subtypes have emerged in human populations as pandemic or epidemic viruses over the last century (H1N1, H3N2 and H2N2). Within some HA subtypes, such as H5, viruses may be further characterised into clades based on genetic differences in their HA, somewhat similar to “variants” in SARS-CoV-2.</p><p>Pathotyping is based on pathogenicity in chickens. Low pathogenicity avian influenza viruses, which are frequently found in wild waterbirds, the natural reservoir for these viruses, generally cause no clinical disease in them or other wild birds. Following introduction into poultry, H5 and H7 avian IAVs may evolve and become highly virulent, causing high levels of mortality in poultry (up to 100% in chickens). Consequently, these viruses are referred to as high pathogenicity avian influenza (HPAI) viruses.</p><p>The ancestor of HPAI H5N1 viruses currently causing a global panzootic (animal pandemic) emerged in geese in China in 1996 (referred to as the Goose/Guandong (gs/gd) lineage) and has since evolved into a continuously circulating group of diverse HPAI viruses. Following a series of key events between 2014 and 2020, a subgroup of the HPAI gs/gd H5N1 lineage (known as clade 2.3.4.4b) emerged and spread like wildfire. Since September 2021, the virus has spread globally, with the exception of Oceania (up to August 2024). The host range has also dramatically increased, with more than 400 wild bird species affected,<span><sup>1</sup></span> and as a result of mass mortality events, some avian taxa may never recover.<span><sup>1</sup></span> This virus is also having a substantial impact on the poultry industry. In 2022, more than 130 000 000 chickens died or were culled because of HPAI.<span><sup>2</sup></span> On top of these large-scale mortality events, an enormous diversity (>50 species) of predatory and scavenging mammalian species have been impacted, ranging from foxes to polar bears to house cats.<span><sup>3</sup></span> For most species, infections have only been sporadic; however, in some instances, unprecedented mass mortality events have occurred, such as in pinnipeds (seal and sea lions) in South America.<span><sup>4</sup></span></p><p>In early 2024 this virus entered dairy cows in the USA by an unknown means and has since infected cows in over 195 herds in 13 states (data as of August 2024).<span><sup>5</sup></span> Although the infection in cows leads to transient mild disease, the infection appears to be concentrated in the udders, leading to very high levels of virus in the milk for 4–6 weeks during infection.<span><sup>6</sup></span> Fortunately, the pasteurisation process destroys the viability of the virus,<span><sup>7</sup></span> but dairy workers are still at risk when milking infected cows as the virus remains infectious on milking equipment, and the unpasteurised milk contains very high virus loads.<span><sup>7</sup></span></p><p>Since 1997, HPAI gs/gd H5N1 has caused nearly 900 human cases in 23 countries, with more than half of these cases resulting in death. To date, the majority of human cases have been those associated with contact with infected poultry, sick backyard birds or linked to poultry markets. However, not all H5N1 cases are caused by the same genetic clade, and differing clades seem to be associated with different clinical outcomes. A prime example is the clade that 2.3.4.4b H5N1 HPAI viruses, which are having a severe effect on wildlife and poultry; however, there have been relatively few human cases reported. Since 2021, globally, there have only been 40 detections of HPAI H5/H5N1. Of these, 17 have been because of clade 2.3.4.4b viruses (Fig. 1). Overall, the Food Agriculture Organization/World Organisation for Animal Health/ World Health Organization (FAO/WOAH/WHO) have indicated in a joint assessment published on 14 August 2024 that ‘the global public health risk of influenza A (H5N1) viruses as low’, but the risk for occupationally exposed people to infection ‘can range from low to medium, depending on the nature of exposure, the duration of exposure and the consistent and appropriate use of personal protective equipment’.<span><sup>8</sup></span> To date, four human cases have been caused by contact with infected dairy cows.<span><sup>9</sup></span> Most human cases infected with 2.3.4.4b H5N1 viruses have been mild<span><sup>10</sup></span> and in some cases may have been because of environmental carriage rather than true infections,<span><sup>11</sup></span> whereas humans who have been infected by cattle have reported mainly conjunctivis.<span><sup>10</sup></span> To date, only a single death has been confirmed caused by a clade 2.3.4.4b HPAI virus.<span><sup>10</sup></span> The Centers for Disease Control and Prevention's (CDC's) Influenza Risk Assessment Tool (IRAT), which uses data from other IAVs, has scored the <i>future</i> pandemic potential of clade 2.3.4.4b as ‘moderate risk’ based on information through 26 June 2024. In a risk comparison between animal influenza strains and subtypes, the 2.3.4.4b currently circulating in cattle has slightly increased in the risk of emergence (but not its impact) compared to other 2.3.4.4b viruses, but its emergence score remained lower than avian H9 and H7N9 viruses found in Asian poultry.<span><sup>12</sup></span></p><p>The ongoing 2.3.4.4b infections across numerous mammalian species are highly concerning, with mammal-to-mammal transmission implicated in outbreaks concerning minks,<span><sup>13</sup></span> pinnipeds<span><sup>4</sup></span> and dairy cows.<span><sup>5</sup></span> Mammal-to-mammal transmission potential has been addressed using ferret experiments. Not only can infection be established in ferrets but it has been demonstrated that these viruses can be transmitted by close contact with naïve ferrets housed in the same cages (and cause disease), but they do not transmit efficiently through aerosols to naïve ferrets separated from the infected animals by a perforated barrier. Four independent studies showed that only four of 18 (22%) ferrets could be infected with different 2.3.4.4b viruses by the aerosol route from infected donors.<span><sup>14-17</sup></span></p><p>For an avian IAV to acquire efficient mammalian transmission through aerosols, several genetic changes would be required. These comprise: (i) changing receptor specificity from avian-type to human-type receptors, i.e. α2.3- to α2.6-linked sialic acids; (ii) increased HA thermostability and pH stability; (iii) increased replication of viruses at temperatures equivalent to those in the mammalian upper respiratory tract; and (iv) changing the expression of RNA transcripts by the viral polymerase complex, resulting in increased replication.</p><p>One of these important changes is a change in the PB2 protein, which is part of the viral replication machinery. A single amino acid change in the PB2 (E627K) causes a change in the polymerase activity, improving the viruses' replication efficiency in mammals.<span><sup>18</sup></span> Interestingly the E627K amino acid change has only rarely been detected in cows infected with the H5N1 in the USA. Other mutations of concern in the PB2 protein which have been shown to increase the polymerase activity of IAVs in mammalian cells or mice, including D701N and Q591K, which were present in viruses detected in pinnipeds in South America<span><sup>4</sup></span>, and T271A (an uncommon mutation) that was identified in a mink H5N1 outbreak in Spain in October 2022.<span><sup>13</sup></span> Higher polymerase activity is likely to lead to higher replication of the virus in a mammalian host. In addition to specific mutations, the mixing of avian IAVs with human or porcine IAVs, might also lead to further mammalian adaptions.<span><sup>19</sup></span> This feature has underpinned all past human pandemic viruses, as all pandemic influenza viruses have contained at least one segment which originated from avian IAV. Fortunately, neither the 2.3.4.4b HPAI viruses nor other HPAI gs/gd H5Nx viruses have found the right combination of mutations or reassortments to make these viruses sufficiently adapted to humans to cause large numbers of human infections, nor have gained the ability to be able to be transmitted efficiently from one infected person to another.</p><p>Globally, it is critical that current, sustained outbreaks in mammals be carefully monitored for the emergence of further mutations, and that these viruses be regularly assessed for their vaccine match, antiviral drug sensitivity, transmission ability in ferret models and binding specificity studies and for pandemic emergence using tools such as IRAT.<span><sup>12</sup></span> In cattle, where control measures are possible, it will be critical to stop spread as continued transmission provides an ‘open air’ experiment for the potential emergence of further mammalian-adapted mutations and spread back into avian populations (which has already occurred) and into other species. Furthermore, H5N1 HPAI viruses for humans must continue to be assessed to ensure that existing candidate vaccine viruses remain well-matched to the circulating strains in the event that vaccines are required. Vaccinations of higher-risk human groups have already begun in some countries, such as Finland.<span><sup>20</sup></span> Effective H5N1 HPAI vaccines for poultry and other livestock may also be useful and could potentially be used to vaccinate threatened, accessible wildlife species.</p><p>Although H5N1 HPAI is not presently in Australia or New Zealand, the risk of arrival with migratory birds is moderate, with high uncertainty. In Australia, nationally coordinated surveillance is undertaken by the National Avian Influenza in Wild Birds group, and response is in accordance with the AUSVETPLAN (https://animalhealthaustralia.com.au/ausvetplan/). Overall, HPAI H5N1 is a complex problem, affecting wildlife, ecosystems, domestic animals, companion animals and humans, and therefore requires a OneHealth response and solution but, fortunately, the risk of this virus progressing to be the cause of the next human pandemic following COVID-19 is currently low.</p>","PeriodicalId":13625,"journal":{"name":"Internal Medicine Journal","volume":"54 11","pages":"1775-1778"},"PeriodicalIF":1.5000,"publicationDate":"2024-10-25","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1111/imj.16550","citationCount":"0","resultStr":"{\"title\":\"The current situation with H5N1 avian influenza and the risk to humans\",\"authors\":\"Michelle Wille, Ian G. Barr\",\"doi\":\"10.1111/imj.16550\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p>Influenza in humans is mainly caused by two different viruses: influenza A virus (IAV) and influenza B virus. Beyond humans, IAV infects a broader range of species, including avian, equine, porcine and canine species. In contrast, influenza B virus almost exclusively infects humans. Avian IAVs are categorised in two main ways: by subtype and by pathotype. Subtyping is based on the diversity of the two major surface proteins: haemagglutinin (HA) and neuraminidase (NA). These proteins are highly diverse, with 19 HA subtypes and 11 NA subtypes, where the HA-NA combination added together make up the basic identity tag, or subtype, for example, H1N1 and H5N1. To date, only three IAV subtypes have emerged in human populations as pandemic or epidemic viruses over the last century (H1N1, H3N2 and H2N2). Within some HA subtypes, such as H5, viruses may be further characterised into clades based on genetic differences in their HA, somewhat similar to “variants” in SARS-CoV-2.</p><p>Pathotyping is based on pathogenicity in chickens. Low pathogenicity avian influenza viruses, which are frequently found in wild waterbirds, the natural reservoir for these viruses, generally cause no clinical disease in them or other wild birds. Following introduction into poultry, H5 and H7 avian IAVs may evolve and become highly virulent, causing high levels of mortality in poultry (up to 100% in chickens). Consequently, these viruses are referred to as high pathogenicity avian influenza (HPAI) viruses.</p><p>The ancestor of HPAI H5N1 viruses currently causing a global panzootic (animal pandemic) emerged in geese in China in 1996 (referred to as the Goose/Guandong (gs/gd) lineage) and has since evolved into a continuously circulating group of diverse HPAI viruses. Following a series of key events between 2014 and 2020, a subgroup of the HPAI gs/gd H5N1 lineage (known as clade 2.3.4.4b) emerged and spread like wildfire. Since September 2021, the virus has spread globally, with the exception of Oceania (up to August 2024). The host range has also dramatically increased, with more than 400 wild bird species affected,<span><sup>1</sup></span> and as a result of mass mortality events, some avian taxa may never recover.<span><sup>1</sup></span> This virus is also having a substantial impact on the poultry industry. In 2022, more than 130 000 000 chickens died or were culled because of HPAI.<span><sup>2</sup></span> On top of these large-scale mortality events, an enormous diversity (>50 species) of predatory and scavenging mammalian species have been impacted, ranging from foxes to polar bears to house cats.<span><sup>3</sup></span> For most species, infections have only been sporadic; however, in some instances, unprecedented mass mortality events have occurred, such as in pinnipeds (seal and sea lions) in South America.<span><sup>4</sup></span></p><p>In early 2024 this virus entered dairy cows in the USA by an unknown means and has since infected cows in over 195 herds in 13 states (data as of August 2024).<span><sup>5</sup></span> Although the infection in cows leads to transient mild disease, the infection appears to be concentrated in the udders, leading to very high levels of virus in the milk for 4–6 weeks during infection.<span><sup>6</sup></span> Fortunately, the pasteurisation process destroys the viability of the virus,<span><sup>7</sup></span> but dairy workers are still at risk when milking infected cows as the virus remains infectious on milking equipment, and the unpasteurised milk contains very high virus loads.<span><sup>7</sup></span></p><p>Since 1997, HPAI gs/gd H5N1 has caused nearly 900 human cases in 23 countries, with more than half of these cases resulting in death. To date, the majority of human cases have been those associated with contact with infected poultry, sick backyard birds or linked to poultry markets. However, not all H5N1 cases are caused by the same genetic clade, and differing clades seem to be associated with different clinical outcomes. A prime example is the clade that 2.3.4.4b H5N1 HPAI viruses, which are having a severe effect on wildlife and poultry; however, there have been relatively few human cases reported. Since 2021, globally, there have only been 40 detections of HPAI H5/H5N1. Of these, 17 have been because of clade 2.3.4.4b viruses (Fig. 1). Overall, the Food Agriculture Organization/World Organisation for Animal Health/ World Health Organization (FAO/WOAH/WHO) have indicated in a joint assessment published on 14 August 2024 that ‘the global public health risk of influenza A (H5N1) viruses as low’, but the risk for occupationally exposed people to infection ‘can range from low to medium, depending on the nature of exposure, the duration of exposure and the consistent and appropriate use of personal protective equipment’.<span><sup>8</sup></span> To date, four human cases have been caused by contact with infected dairy cows.<span><sup>9</sup></span> Most human cases infected with 2.3.4.4b H5N1 viruses have been mild<span><sup>10</sup></span> and in some cases may have been because of environmental carriage rather than true infections,<span><sup>11</sup></span> whereas humans who have been infected by cattle have reported mainly conjunctivis.<span><sup>10</sup></span> To date, only a single death has been confirmed caused by a clade 2.3.4.4b HPAI virus.<span><sup>10</sup></span> The Centers for Disease Control and Prevention's (CDC's) Influenza Risk Assessment Tool (IRAT), which uses data from other IAVs, has scored the <i>future</i> pandemic potential of clade 2.3.4.4b as ‘moderate risk’ based on information through 26 June 2024. In a risk comparison between animal influenza strains and subtypes, the 2.3.4.4b currently circulating in cattle has slightly increased in the risk of emergence (but not its impact) compared to other 2.3.4.4b viruses, but its emergence score remained lower than avian H9 and H7N9 viruses found in Asian poultry.<span><sup>12</sup></span></p><p>The ongoing 2.3.4.4b infections across numerous mammalian species are highly concerning, with mammal-to-mammal transmission implicated in outbreaks concerning minks,<span><sup>13</sup></span> pinnipeds<span><sup>4</sup></span> and dairy cows.<span><sup>5</sup></span> Mammal-to-mammal transmission potential has been addressed using ferret experiments. Not only can infection be established in ferrets but it has been demonstrated that these viruses can be transmitted by close contact with naïve ferrets housed in the same cages (and cause disease), but they do not transmit efficiently through aerosols to naïve ferrets separated from the infected animals by a perforated barrier. Four independent studies showed that only four of 18 (22%) ferrets could be infected with different 2.3.4.4b viruses by the aerosol route from infected donors.<span><sup>14-17</sup></span></p><p>For an avian IAV to acquire efficient mammalian transmission through aerosols, several genetic changes would be required. These comprise: (i) changing receptor specificity from avian-type to human-type receptors, i.e. α2.3- to α2.6-linked sialic acids; (ii) increased HA thermostability and pH stability; (iii) increased replication of viruses at temperatures equivalent to those in the mammalian upper respiratory tract; and (iv) changing the expression of RNA transcripts by the viral polymerase complex, resulting in increased replication.</p><p>One of these important changes is a change in the PB2 protein, which is part of the viral replication machinery. A single amino acid change in the PB2 (E627K) causes a change in the polymerase activity, improving the viruses' replication efficiency in mammals.<span><sup>18</sup></span> Interestingly the E627K amino acid change has only rarely been detected in cows infected with the H5N1 in the USA. Other mutations of concern in the PB2 protein which have been shown to increase the polymerase activity of IAVs in mammalian cells or mice, including D701N and Q591K, which were present in viruses detected in pinnipeds in South America<span><sup>4</sup></span>, and T271A (an uncommon mutation) that was identified in a mink H5N1 outbreak in Spain in October 2022.<span><sup>13</sup></span> Higher polymerase activity is likely to lead to higher replication of the virus in a mammalian host. In addition to specific mutations, the mixing of avian IAVs with human or porcine IAVs, might also lead to further mammalian adaptions.<span><sup>19</sup></span> This feature has underpinned all past human pandemic viruses, as all pandemic influenza viruses have contained at least one segment which originated from avian IAV. Fortunately, neither the 2.3.4.4b HPAI viruses nor other HPAI gs/gd H5Nx viruses have found the right combination of mutations or reassortments to make these viruses sufficiently adapted to humans to cause large numbers of human infections, nor have gained the ability to be able to be transmitted efficiently from one infected person to another.</p><p>Globally, it is critical that current, sustained outbreaks in mammals be carefully monitored for the emergence of further mutations, and that these viruses be regularly assessed for their vaccine match, antiviral drug sensitivity, transmission ability in ferret models and binding specificity studies and for pandemic emergence using tools such as IRAT.<span><sup>12</sup></span> In cattle, where control measures are possible, it will be critical to stop spread as continued transmission provides an ‘open air’ experiment for the potential emergence of further mammalian-adapted mutations and spread back into avian populations (which has already occurred) and into other species. Furthermore, H5N1 HPAI viruses for humans must continue to be assessed to ensure that existing candidate vaccine viruses remain well-matched to the circulating strains in the event that vaccines are required. Vaccinations of higher-risk human groups have already begun in some countries, such as Finland.<span><sup>20</sup></span> Effective H5N1 HPAI vaccines for poultry and other livestock may also be useful and could potentially be used to vaccinate threatened, accessible wildlife species.</p><p>Although H5N1 HPAI is not presently in Australia or New Zealand, the risk of arrival with migratory birds is moderate, with high uncertainty. In Australia, nationally coordinated surveillance is undertaken by the National Avian Influenza in Wild Birds group, and response is in accordance with the AUSVETPLAN (https://animalhealthaustralia.com.au/ausvetplan/). Overall, HPAI H5N1 is a complex problem, affecting wildlife, ecosystems, domestic animals, companion animals and humans, and therefore requires a OneHealth response and solution but, fortunately, the risk of this virus progressing to be the cause of the next human pandemic following COVID-19 is currently low.</p>\",\"PeriodicalId\":13625,\"journal\":{\"name\":\"Internal Medicine Journal\",\"volume\":\"54 11\",\"pages\":\"1775-1778\"},\"PeriodicalIF\":1.5000,\"publicationDate\":\"2024-10-25\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://onlinelibrary.wiley.com/doi/epdf/10.1111/imj.16550\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"Internal Medicine Journal\",\"FirstCategoryId\":\"3\",\"ListUrlMain\":\"https://onlinelibrary.wiley.com/doi/10.1111/imj.16550\",\"RegionNum\":4,\"RegionCategory\":\"医学\",\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"Q2\",\"JCRName\":\"MEDICINE, GENERAL & INTERNAL\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"Internal Medicine Journal","FirstCategoryId":"3","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1111/imj.16550","RegionNum":4,"RegionCategory":"医学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q2","JCRName":"MEDICINE, GENERAL & INTERNAL","Score":null,"Total":0}
The current situation with H5N1 avian influenza and the risk to humans
Influenza in humans is mainly caused by two different viruses: influenza A virus (IAV) and influenza B virus. Beyond humans, IAV infects a broader range of species, including avian, equine, porcine and canine species. In contrast, influenza B virus almost exclusively infects humans. Avian IAVs are categorised in two main ways: by subtype and by pathotype. Subtyping is based on the diversity of the two major surface proteins: haemagglutinin (HA) and neuraminidase (NA). These proteins are highly diverse, with 19 HA subtypes and 11 NA subtypes, where the HA-NA combination added together make up the basic identity tag, or subtype, for example, H1N1 and H5N1. To date, only three IAV subtypes have emerged in human populations as pandemic or epidemic viruses over the last century (H1N1, H3N2 and H2N2). Within some HA subtypes, such as H5, viruses may be further characterised into clades based on genetic differences in their HA, somewhat similar to “variants” in SARS-CoV-2.
Pathotyping is based on pathogenicity in chickens. Low pathogenicity avian influenza viruses, which are frequently found in wild waterbirds, the natural reservoir for these viruses, generally cause no clinical disease in them or other wild birds. Following introduction into poultry, H5 and H7 avian IAVs may evolve and become highly virulent, causing high levels of mortality in poultry (up to 100% in chickens). Consequently, these viruses are referred to as high pathogenicity avian influenza (HPAI) viruses.
The ancestor of HPAI H5N1 viruses currently causing a global panzootic (animal pandemic) emerged in geese in China in 1996 (referred to as the Goose/Guandong (gs/gd) lineage) and has since evolved into a continuously circulating group of diverse HPAI viruses. Following a series of key events between 2014 and 2020, a subgroup of the HPAI gs/gd H5N1 lineage (known as clade 2.3.4.4b) emerged and spread like wildfire. Since September 2021, the virus has spread globally, with the exception of Oceania (up to August 2024). The host range has also dramatically increased, with more than 400 wild bird species affected,1 and as a result of mass mortality events, some avian taxa may never recover.1 This virus is also having a substantial impact on the poultry industry. In 2022, more than 130 000 000 chickens died or were culled because of HPAI.2 On top of these large-scale mortality events, an enormous diversity (>50 species) of predatory and scavenging mammalian species have been impacted, ranging from foxes to polar bears to house cats.3 For most species, infections have only been sporadic; however, in some instances, unprecedented mass mortality events have occurred, such as in pinnipeds (seal and sea lions) in South America.4
In early 2024 this virus entered dairy cows in the USA by an unknown means and has since infected cows in over 195 herds in 13 states (data as of August 2024).5 Although the infection in cows leads to transient mild disease, the infection appears to be concentrated in the udders, leading to very high levels of virus in the milk for 4–6 weeks during infection.6 Fortunately, the pasteurisation process destroys the viability of the virus,7 but dairy workers are still at risk when milking infected cows as the virus remains infectious on milking equipment, and the unpasteurised milk contains very high virus loads.7
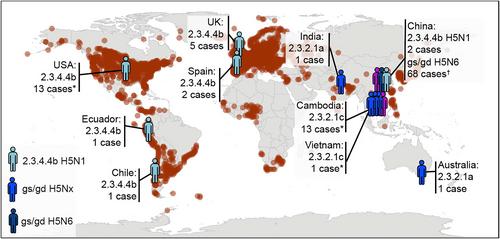
Since 1997, HPAI gs/gd H5N1 has caused nearly 900 human cases in 23 countries, with more than half of these cases resulting in death. To date, the majority of human cases have been those associated with contact with infected poultry, sick backyard birds or linked to poultry markets. However, not all H5N1 cases are caused by the same genetic clade, and differing clades seem to be associated with different clinical outcomes. A prime example is the clade that 2.3.4.4b H5N1 HPAI viruses, which are having a severe effect on wildlife and poultry; however, there have been relatively few human cases reported. Since 2021, globally, there have only been 40 detections of HPAI H5/H5N1. Of these, 17 have been because of clade 2.3.4.4b viruses (Fig. 1). Overall, the Food Agriculture Organization/World Organisation for Animal Health/ World Health Organization (FAO/WOAH/WHO) have indicated in a joint assessment published on 14 August 2024 that ‘the global public health risk of influenza A (H5N1) viruses as low’, but the risk for occupationally exposed people to infection ‘can range from low to medium, depending on the nature of exposure, the duration of exposure and the consistent and appropriate use of personal protective equipment’.8 To date, four human cases have been caused by contact with infected dairy cows.9 Most human cases infected with 2.3.4.4b H5N1 viruses have been mild10 and in some cases may have been because of environmental carriage rather than true infections,11 whereas humans who have been infected by cattle have reported mainly conjunctivis.10 To date, only a single death has been confirmed caused by a clade 2.3.4.4b HPAI virus.10 The Centers for Disease Control and Prevention's (CDC's) Influenza Risk Assessment Tool (IRAT), which uses data from other IAVs, has scored the future pandemic potential of clade 2.3.4.4b as ‘moderate risk’ based on information through 26 June 2024. In a risk comparison between animal influenza strains and subtypes, the 2.3.4.4b currently circulating in cattle has slightly increased in the risk of emergence (but not its impact) compared to other 2.3.4.4b viruses, but its emergence score remained lower than avian H9 and H7N9 viruses found in Asian poultry.12
The ongoing 2.3.4.4b infections across numerous mammalian species are highly concerning, with mammal-to-mammal transmission implicated in outbreaks concerning minks,13 pinnipeds4 and dairy cows.5 Mammal-to-mammal transmission potential has been addressed using ferret experiments. Not only can infection be established in ferrets but it has been demonstrated that these viruses can be transmitted by close contact with naïve ferrets housed in the same cages (and cause disease), but they do not transmit efficiently through aerosols to naïve ferrets separated from the infected animals by a perforated barrier. Four independent studies showed that only four of 18 (22%) ferrets could be infected with different 2.3.4.4b viruses by the aerosol route from infected donors.14-17
For an avian IAV to acquire efficient mammalian transmission through aerosols, several genetic changes would be required. These comprise: (i) changing receptor specificity from avian-type to human-type receptors, i.e. α2.3- to α2.6-linked sialic acids; (ii) increased HA thermostability and pH stability; (iii) increased replication of viruses at temperatures equivalent to those in the mammalian upper respiratory tract; and (iv) changing the expression of RNA transcripts by the viral polymerase complex, resulting in increased replication.
One of these important changes is a change in the PB2 protein, which is part of the viral replication machinery. A single amino acid change in the PB2 (E627K) causes a change in the polymerase activity, improving the viruses' replication efficiency in mammals.18 Interestingly the E627K amino acid change has only rarely been detected in cows infected with the H5N1 in the USA. Other mutations of concern in the PB2 protein which have been shown to increase the polymerase activity of IAVs in mammalian cells or mice, including D701N and Q591K, which were present in viruses detected in pinnipeds in South America4, and T271A (an uncommon mutation) that was identified in a mink H5N1 outbreak in Spain in October 2022.13 Higher polymerase activity is likely to lead to higher replication of the virus in a mammalian host. In addition to specific mutations, the mixing of avian IAVs with human or porcine IAVs, might also lead to further mammalian adaptions.19 This feature has underpinned all past human pandemic viruses, as all pandemic influenza viruses have contained at least one segment which originated from avian IAV. Fortunately, neither the 2.3.4.4b HPAI viruses nor other HPAI gs/gd H5Nx viruses have found the right combination of mutations or reassortments to make these viruses sufficiently adapted to humans to cause large numbers of human infections, nor have gained the ability to be able to be transmitted efficiently from one infected person to another.
Globally, it is critical that current, sustained outbreaks in mammals be carefully monitored for the emergence of further mutations, and that these viruses be regularly assessed for their vaccine match, antiviral drug sensitivity, transmission ability in ferret models and binding specificity studies and for pandemic emergence using tools such as IRAT.12 In cattle, where control measures are possible, it will be critical to stop spread as continued transmission provides an ‘open air’ experiment for the potential emergence of further mammalian-adapted mutations and spread back into avian populations (which has already occurred) and into other species. Furthermore, H5N1 HPAI viruses for humans must continue to be assessed to ensure that existing candidate vaccine viruses remain well-matched to the circulating strains in the event that vaccines are required. Vaccinations of higher-risk human groups have already begun in some countries, such as Finland.20 Effective H5N1 HPAI vaccines for poultry and other livestock may also be useful and could potentially be used to vaccinate threatened, accessible wildlife species.
Although H5N1 HPAI is not presently in Australia or New Zealand, the risk of arrival with migratory birds is moderate, with high uncertainty. In Australia, nationally coordinated surveillance is undertaken by the National Avian Influenza in Wild Birds group, and response is in accordance with the AUSVETPLAN (https://animalhealthaustralia.com.au/ausvetplan/). Overall, HPAI H5N1 is a complex problem, affecting wildlife, ecosystems, domestic animals, companion animals and humans, and therefore requires a OneHealth response and solution but, fortunately, the risk of this virus progressing to be the cause of the next human pandemic following COVID-19 is currently low.
期刊介绍:
The Internal Medicine Journal is the official journal of the Adult Medicine Division of The Royal Australasian College of Physicians (RACP). Its purpose is to publish high-quality internationally competitive peer-reviewed original medical research, both laboratory and clinical, relating to the study and research of human disease. Papers will be considered from all areas of medical practice and science. The Journal also has a major role in continuing medical education and publishes review articles relevant to physician education.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


