Yuan Wang, Maria-Filothei Lazaridou, Theresa Kordaß, Chiara Massa, Christoforos K. Vaxevanis, Stefan Eichmüller, Barbara Seliger
{"title":"非常规 microRNA 作用:通过与沉默器相互作用增强人类白细胞抗原 I 类抗原处理途径。","authors":"Yuan Wang, Maria-Filothei Lazaridou, Theresa Kordaß, Chiara Massa, Christoforos K. Vaxevanis, Stefan Eichmüller, Barbara Seliger","doi":"10.1002/ctm2.70010","DOIUrl":null,"url":null,"abstract":"<p>Dear Editor,</p><p>The unconventional functional mechanisms of microRNA (miRNA)-mediated RNA or protein activation are complex and diverse,<span><sup>1</sup></span> like miRNA binding to AU-rich elements (ARE)<span><sup>2</sup></span> or competing with RNA-binding proteins.<span><sup>3</sup></span> Our own data demonstrated that miR-16 could bind to the coding sequence (CDS) of classical and non-classical human leukocyte antigen class I (HLA-I) molecules, thereby inducing their expression.<span><sup>4</sup></span> However, the interaction of silencers with miRNAs has not yet been investigated. Silencer features include a high GC content,<span><sup>5</sup></span> DNase hypersensitivity sites<span><sup>6</sup></span> and H3K27me3 regions.<span><sup>7</sup></span> Here we identified for the first time that miR-155-5p can directly bind a silencer in the 3′untranslated region (3′UTR) of TAP-binding protein (tpn) thereby increasing the HLA-I surface expression.</p><p>Using miRNA trapping by RNA in vitro affinity purification (miTRAP),<span><sup>8</sup></span> in silico analyses and molecular experiments, we identified miR-155-5p targeting of tpn 3′ UTR in melanoma cells affecting tpn and cell surface HLA-I expression, which has also clinical relevance. Interestingly, upon deletion of the predicted binding site within tpn 3´UTR (Figure 1A), luciferase (luc) reporter assays indicated higher relative luc activity of the wild type (wt) compared to the del 3′UTR in HEK293T cells (Figure 1B), which is opposite to the conventional function of miRNAs leading to a downregulation. Overexpression of miR-155-5p in three melanoma cell lines (Figure 1C) increased their tpn messenger RNA (mRNA) (Figure 1D) and protein levels (Figure 1E,F). This upregulation was specific for tpn since the mRNA of programmed death ligand 1 (PD-L1), another target of miR-155-5p,<span><sup>9</sup></span> was downregulated in the miR-155-5p-transfected MZ-Mel2 cell line (Figure 1G). Despite the overall expression levels of the HLA-I heavy chain (HC) were not altered in the miR-155-5p transfectants (Figure 1F,H), a tpn-mediated upregulation of the HLA-ABC and HLA-BC surface antigens was found on FM81 and MZ-Mel2 cells (Figure 1I,J), but not on FM3 cells, which might be probably due to the high HLA-I surface expression when compared to FM81 and MZ-Mel2 cells (Figure 1F). Actinomycin D treatment revealed a significant increase in the tpn mRNA half-life in FM81 miR-155-5p transfectants (Figure 1K). Using a CD107a degranulation assay, a reduced NK cell-mediated cytotoxicity against miR-155-5p transfected MZ-Mel2 cells expressing increased HLA-I surface antigens was shown by lower numbers of CD107a-positive NK cells (Figure 1L).</p><p>The overall survival (OS) analysis of 214 metastatic melanoma cases with patients’ outcomes demonstrated a positive correlation of miR-155 (miR-155HG) (Figure 2A), tpn (Figure 2B) and HLA-A (Figure 2C) expression levels with the OS of patients, thereby confirming ours in vitro experiments. Similar results were obtained by bioinformatics analyses of all 444 cases or 63 cases of distant metastatic melanoma from the “TCGA Skin Cutaneous Melanoma (SKCM)” dataset (Figure 2D,E). Furthermore, a strong positive correlation between miR-155 expression and the frequency of CD8<sup>+</sup> T cells was found in this dataset using the CIBERSORT web tool (Figure 2F,G).</p><p>Subsequently, in silico analysis of the miRNA binding site sequence in the 3′ UTR of tpn revealed a high GC content and a DNase hypersensitivity site (DHS) suggesting its role as a part of a silencer (Table S1 and Figure S1). Three sequences (sil1, sil3, sil4) upstream of the binding site, an AU-rich element predicted via the ARE site web tool and the potential GA-, AU- and GC-rich areas within the miR-155-5p binding site were deleted in the tpn 3´UTR (Figure 3A and Figure S2A) and cloned into the miR-GLO-vector since the miRNA-mediated activation<span><sup>10</sup></span> is linked to the binding to specific sequence elements. The luc reporter assay revealed that in comparison to the wt 3′UTR, the transfection of AREdel, AUdel, GCdel, or GAdel vectors into the HEK293T cells reverted the suppressive effects in the negative control (NC), parental cells and the positive effect in miR-155-5p transfectants. Since these elements belong to the miR-155-5p binding site, their deletions have disrupted the binding site leading to the loss of the positive miR-155-5p function on tpn after transfection confirming these elements represent parts of the miR-155-5p binding site and the core part of a silencer. Whereas in both NC and parental cells, the repressive effects of sil1del, sil3del or sil4del groups were similar to the wt group, all sil groups in the miR-155-5p transfected cells have positive, but much lower effects than in the wt group (Figure 3B) suggesting the disruption of secondary mRNA structures by the partial sequence deletions (Figure S2B–K) are involved in this enhancement.</p><p>In addition, a CRISPR/Cas9-mediated deletion of the miR-155-5p binding site was generated in the FM3 melanoma cell line upon transfection of modified PX458 (Figure 3C and Figure S2A). Sanger sequencing (data not shown) and PCR amplification (Figure S2L) demonstrated the successful deletion of the miR-155-5p binding site (R3R9) resulting in an upregulation of tpn mRNA and protein levels when compared to the mock vector (PX458) (Figure 3D–F). As shown in Figure 3G, the tpn expression (green) was higher in the miR-155 group than in the other two groups, while the expression of H3K27me3 (red) was inversely correlated due to direct or indirect binding of tpn to H3K27me3 as shown by immunoprecipitation (Figure S2M), which strengthen the evidence that the binding site is part of a silencer.</p><p>Finally, to exclude that the miRNA-mediated activation of tpn was caused by RBPs, the four proteins HNRNPL, HNRNPC, IGF2BP1 and IGF2BP3, known to bind to the tpn 3′UTR, were explored. Compared to HNRNPC and HNRNPL, the IGF2BP1 and IGF2BP3 binding sites overlap with that of miR-155-5p (Figure S3A–D). Interestingly, the miR-155-5p binding site is a part of the IGF2BP1 and IGF2BP3 potential binding sites containing a GC- and an AU-rich element (Figure S3E). Concentration gradient silencing assays revealed an interaction of miR-155-5p and these RBPs (Figure S3F–H).</p><p>This study proposes a new unconventional function of miRNAs that enhances target transcription through binding a silencer thereby activating the downstream pathway(s) (Figure 4). Using miR-155-5p as a model, this is the first report (i) identifying a silencer in the tpn 3′UTR, which (ii) directly interacts with a non-coding RNA and (iii) has clinical relevance. These data extend miRNAs’ functions and add new insights to our knowledge of silencers.</p><p>Conceptualization: Yuan Wang, Barbara Seliger, Theresa Kordaß and Stefan Eichmüller. Methodology: Yuan Wang, Maria-Filothei Lazaridou, Chiara Massa, Theresa Kordaß, Christoforos K. Vaxevanis. Investigation: Yuan Wang, Maria-Filothei Lazaridou and Chiara Massa. Visualization: Yuan Wang, Maria-Filothei Lazaridou and Chiara Massa. Funding acquisition: Barbara Seliger and Stefan Eichmüller; Writing—original draft: Yuan Wang and Barbara Seliger; Writing—review & editing: Yuan Wang, Barbara Seliger, Theresa Kordaß, Stefan Eichmüller, Maria-Filothei Lazaridou, Chiara Massa, Christoforos K. Vaxevanis.</p><p>The authors declare no conflict of interest.</p><p>The work was funded by grants of the Deutsche Krebshilfe, grant 341025929 (Barbara Seliger) and the Deutsche Forschungsgemeinschaft, grant number: SE581/33-1.</p>","PeriodicalId":10189,"journal":{"name":"Clinical and Translational Medicine","volume":null,"pages":null},"PeriodicalIF":7.9000,"publicationDate":"2024-10-22","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://www.ncbi.nlm.nih.gov/pmc/articles/PMC11496566/pdf/","citationCount":"0","resultStr":"{\"title\":\"Unconventional microRNA role: Enhancing the human leukocyte antigen class I antigen processing pathway via interacting with a silencer\",\"authors\":\"Yuan Wang, Maria-Filothei Lazaridou, Theresa Kordaß, Chiara Massa, Christoforos K. Vaxevanis, Stefan Eichmüller, Barbara Seliger\",\"doi\":\"10.1002/ctm2.70010\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p>Dear Editor,</p><p>The unconventional functional mechanisms of microRNA (miRNA)-mediated RNA or protein activation are complex and diverse,<span><sup>1</sup></span> like miRNA binding to AU-rich elements (ARE)<span><sup>2</sup></span> or competing with RNA-binding proteins.<span><sup>3</sup></span> Our own data demonstrated that miR-16 could bind to the coding sequence (CDS) of classical and non-classical human leukocyte antigen class I (HLA-I) molecules, thereby inducing their expression.<span><sup>4</sup></span> However, the interaction of silencers with miRNAs has not yet been investigated. Silencer features include a high GC content,<span><sup>5</sup></span> DNase hypersensitivity sites<span><sup>6</sup></span> and H3K27me3 regions.<span><sup>7</sup></span> Here we identified for the first time that miR-155-5p can directly bind a silencer in the 3′untranslated region (3′UTR) of TAP-binding protein (tpn) thereby increasing the HLA-I surface expression.</p><p>Using miRNA trapping by RNA in vitro affinity purification (miTRAP),<span><sup>8</sup></span> in silico analyses and molecular experiments, we identified miR-155-5p targeting of tpn 3′ UTR in melanoma cells affecting tpn and cell surface HLA-I expression, which has also clinical relevance. Interestingly, upon deletion of the predicted binding site within tpn 3´UTR (Figure 1A), luciferase (luc) reporter assays indicated higher relative luc activity of the wild type (wt) compared to the del 3′UTR in HEK293T cells (Figure 1B), which is opposite to the conventional function of miRNAs leading to a downregulation. Overexpression of miR-155-5p in three melanoma cell lines (Figure 1C) increased their tpn messenger RNA (mRNA) (Figure 1D) and protein levels (Figure 1E,F). This upregulation was specific for tpn since the mRNA of programmed death ligand 1 (PD-L1), another target of miR-155-5p,<span><sup>9</sup></span> was downregulated in the miR-155-5p-transfected MZ-Mel2 cell line (Figure 1G). Despite the overall expression levels of the HLA-I heavy chain (HC) were not altered in the miR-155-5p transfectants (Figure 1F,H), a tpn-mediated upregulation of the HLA-ABC and HLA-BC surface antigens was found on FM81 and MZ-Mel2 cells (Figure 1I,J), but not on FM3 cells, which might be probably due to the high HLA-I surface expression when compared to FM81 and MZ-Mel2 cells (Figure 1F). Actinomycin D treatment revealed a significant increase in the tpn mRNA half-life in FM81 miR-155-5p transfectants (Figure 1K). Using a CD107a degranulation assay, a reduced NK cell-mediated cytotoxicity against miR-155-5p transfected MZ-Mel2 cells expressing increased HLA-I surface antigens was shown by lower numbers of CD107a-positive NK cells (Figure 1L).</p><p>The overall survival (OS) analysis of 214 metastatic melanoma cases with patients’ outcomes demonstrated a positive correlation of miR-155 (miR-155HG) (Figure 2A), tpn (Figure 2B) and HLA-A (Figure 2C) expression levels with the OS of patients, thereby confirming ours in vitro experiments. Similar results were obtained by bioinformatics analyses of all 444 cases or 63 cases of distant metastatic melanoma from the “TCGA Skin Cutaneous Melanoma (SKCM)” dataset (Figure 2D,E). Furthermore, a strong positive correlation between miR-155 expression and the frequency of CD8<sup>+</sup> T cells was found in this dataset using the CIBERSORT web tool (Figure 2F,G).</p><p>Subsequently, in silico analysis of the miRNA binding site sequence in the 3′ UTR of tpn revealed a high GC content and a DNase hypersensitivity site (DHS) suggesting its role as a part of a silencer (Table S1 and Figure S1). Three sequences (sil1, sil3, sil4) upstream of the binding site, an AU-rich element predicted via the ARE site web tool and the potential GA-, AU- and GC-rich areas within the miR-155-5p binding site were deleted in the tpn 3´UTR (Figure 3A and Figure S2A) and cloned into the miR-GLO-vector since the miRNA-mediated activation<span><sup>10</sup></span> is linked to the binding to specific sequence elements. The luc reporter assay revealed that in comparison to the wt 3′UTR, the transfection of AREdel, AUdel, GCdel, or GAdel vectors into the HEK293T cells reverted the suppressive effects in the negative control (NC), parental cells and the positive effect in miR-155-5p transfectants. Since these elements belong to the miR-155-5p binding site, their deletions have disrupted the binding site leading to the loss of the positive miR-155-5p function on tpn after transfection confirming these elements represent parts of the miR-155-5p binding site and the core part of a silencer. Whereas in both NC and parental cells, the repressive effects of sil1del, sil3del or sil4del groups were similar to the wt group, all sil groups in the miR-155-5p transfected cells have positive, but much lower effects than in the wt group (Figure 3B) suggesting the disruption of secondary mRNA structures by the partial sequence deletions (Figure S2B–K) are involved in this enhancement.</p><p>In addition, a CRISPR/Cas9-mediated deletion of the miR-155-5p binding site was generated in the FM3 melanoma cell line upon transfection of modified PX458 (Figure 3C and Figure S2A). Sanger sequencing (data not shown) and PCR amplification (Figure S2L) demonstrated the successful deletion of the miR-155-5p binding site (R3R9) resulting in an upregulation of tpn mRNA and protein levels when compared to the mock vector (PX458) (Figure 3D–F). As shown in Figure 3G, the tpn expression (green) was higher in the miR-155 group than in the other two groups, while the expression of H3K27me3 (red) was inversely correlated due to direct or indirect binding of tpn to H3K27me3 as shown by immunoprecipitation (Figure S2M), which strengthen the evidence that the binding site is part of a silencer.</p><p>Finally, to exclude that the miRNA-mediated activation of tpn was caused by RBPs, the four proteins HNRNPL, HNRNPC, IGF2BP1 and IGF2BP3, known to bind to the tpn 3′UTR, were explored. Compared to HNRNPC and HNRNPL, the IGF2BP1 and IGF2BP3 binding sites overlap with that of miR-155-5p (Figure S3A–D). Interestingly, the miR-155-5p binding site is a part of the IGF2BP1 and IGF2BP3 potential binding sites containing a GC- and an AU-rich element (Figure S3E). Concentration gradient silencing assays revealed an interaction of miR-155-5p and these RBPs (Figure S3F–H).</p><p>This study proposes a new unconventional function of miRNAs that enhances target transcription through binding a silencer thereby activating the downstream pathway(s) (Figure 4). Using miR-155-5p as a model, this is the first report (i) identifying a silencer in the tpn 3′UTR, which (ii) directly interacts with a non-coding RNA and (iii) has clinical relevance. These data extend miRNAs’ functions and add new insights to our knowledge of silencers.</p><p>Conceptualization: Yuan Wang, Barbara Seliger, Theresa Kordaß and Stefan Eichmüller. Methodology: Yuan Wang, Maria-Filothei Lazaridou, Chiara Massa, Theresa Kordaß, Christoforos K. Vaxevanis. Investigation: Yuan Wang, Maria-Filothei Lazaridou and Chiara Massa. Visualization: Yuan Wang, Maria-Filothei Lazaridou and Chiara Massa. Funding acquisition: Barbara Seliger and Stefan Eichmüller; Writing—original draft: Yuan Wang and Barbara Seliger; Writing—review & editing: Yuan Wang, Barbara Seliger, Theresa Kordaß, Stefan Eichmüller, Maria-Filothei Lazaridou, Chiara Massa, Christoforos K. Vaxevanis.</p><p>The authors declare no conflict of interest.</p><p>The work was funded by grants of the Deutsche Krebshilfe, grant 341025929 (Barbara Seliger) and the Deutsche Forschungsgemeinschaft, grant number: SE581/33-1.</p>\",\"PeriodicalId\":10189,\"journal\":{\"name\":\"Clinical and Translational Medicine\",\"volume\":null,\"pages\":null},\"PeriodicalIF\":7.9000,\"publicationDate\":\"2024-10-22\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://www.ncbi.nlm.nih.gov/pmc/articles/PMC11496566/pdf/\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"Clinical and Translational Medicine\",\"FirstCategoryId\":\"3\",\"ListUrlMain\":\"https://onlinelibrary.wiley.com/doi/10.1002/ctm2.70010\",\"RegionNum\":1,\"RegionCategory\":\"医学\",\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"Q1\",\"JCRName\":\"MEDICINE, RESEARCH & EXPERIMENTAL\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"Clinical and Translational Medicine","FirstCategoryId":"3","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1002/ctm2.70010","RegionNum":1,"RegionCategory":"医学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"MEDICINE, RESEARCH & EXPERIMENTAL","Score":null,"Total":0}
Unconventional microRNA role: Enhancing the human leukocyte antigen class I antigen processing pathway via interacting with a silencer
Dear Editor,
The unconventional functional mechanisms of microRNA (miRNA)-mediated RNA or protein activation are complex and diverse,1 like miRNA binding to AU-rich elements (ARE)2 or competing with RNA-binding proteins.3 Our own data demonstrated that miR-16 could bind to the coding sequence (CDS) of classical and non-classical human leukocyte antigen class I (HLA-I) molecules, thereby inducing their expression.4 However, the interaction of silencers with miRNAs has not yet been investigated. Silencer features include a high GC content,5 DNase hypersensitivity sites6 and H3K27me3 regions.7 Here we identified for the first time that miR-155-5p can directly bind a silencer in the 3′untranslated region (3′UTR) of TAP-binding protein (tpn) thereby increasing the HLA-I surface expression.
Using miRNA trapping by RNA in vitro affinity purification (miTRAP),8 in silico analyses and molecular experiments, we identified miR-155-5p targeting of tpn 3′ UTR in melanoma cells affecting tpn and cell surface HLA-I expression, which has also clinical relevance. Interestingly, upon deletion of the predicted binding site within tpn 3´UTR (Figure 1A), luciferase (luc) reporter assays indicated higher relative luc activity of the wild type (wt) compared to the del 3′UTR in HEK293T cells (Figure 1B), which is opposite to the conventional function of miRNAs leading to a downregulation. Overexpression of miR-155-5p in three melanoma cell lines (Figure 1C) increased their tpn messenger RNA (mRNA) (Figure 1D) and protein levels (Figure 1E,F). This upregulation was specific for tpn since the mRNA of programmed death ligand 1 (PD-L1), another target of miR-155-5p,9 was downregulated in the miR-155-5p-transfected MZ-Mel2 cell line (Figure 1G). Despite the overall expression levels of the HLA-I heavy chain (HC) were not altered in the miR-155-5p transfectants (Figure 1F,H), a tpn-mediated upregulation of the HLA-ABC and HLA-BC surface antigens was found on FM81 and MZ-Mel2 cells (Figure 1I,J), but not on FM3 cells, which might be probably due to the high HLA-I surface expression when compared to FM81 and MZ-Mel2 cells (Figure 1F). Actinomycin D treatment revealed a significant increase in the tpn mRNA half-life in FM81 miR-155-5p transfectants (Figure 1K). Using a CD107a degranulation assay, a reduced NK cell-mediated cytotoxicity against miR-155-5p transfected MZ-Mel2 cells expressing increased HLA-I surface antigens was shown by lower numbers of CD107a-positive NK cells (Figure 1L).
The overall survival (OS) analysis of 214 metastatic melanoma cases with patients’ outcomes demonstrated a positive correlation of miR-155 (miR-155HG) (Figure 2A), tpn (Figure 2B) and HLA-A (Figure 2C) expression levels with the OS of patients, thereby confirming ours in vitro experiments. Similar results were obtained by bioinformatics analyses of all 444 cases or 63 cases of distant metastatic melanoma from the “TCGA Skin Cutaneous Melanoma (SKCM)” dataset (Figure 2D,E). Furthermore, a strong positive correlation between miR-155 expression and the frequency of CD8+ T cells was found in this dataset using the CIBERSORT web tool (Figure 2F,G).
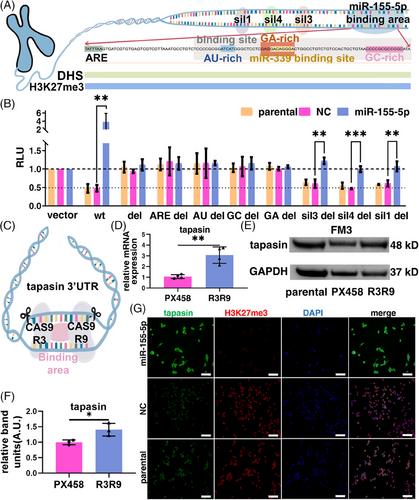
Subsequently, in silico analysis of the miRNA binding site sequence in the 3′ UTR of tpn revealed a high GC content and a DNase hypersensitivity site (DHS) suggesting its role as a part of a silencer (Table S1 and Figure S1). Three sequences (sil1, sil3, sil4) upstream of the binding site, an AU-rich element predicted via the ARE site web tool and the potential GA-, AU- and GC-rich areas within the miR-155-5p binding site were deleted in the tpn 3´UTR (Figure 3A and Figure S2A) and cloned into the miR-GLO-vector since the miRNA-mediated activation10 is linked to the binding to specific sequence elements. The luc reporter assay revealed that in comparison to the wt 3′UTR, the transfection of AREdel, AUdel, GCdel, or GAdel vectors into the HEK293T cells reverted the suppressive effects in the negative control (NC), parental cells and the positive effect in miR-155-5p transfectants. Since these elements belong to the miR-155-5p binding site, their deletions have disrupted the binding site leading to the loss of the positive miR-155-5p function on tpn after transfection confirming these elements represent parts of the miR-155-5p binding site and the core part of a silencer. Whereas in both NC and parental cells, the repressive effects of sil1del, sil3del or sil4del groups were similar to the wt group, all sil groups in the miR-155-5p transfected cells have positive, but much lower effects than in the wt group (Figure 3B) suggesting the disruption of secondary mRNA structures by the partial sequence deletions (Figure S2B–K) are involved in this enhancement.
In addition, a CRISPR/Cas9-mediated deletion of the miR-155-5p binding site was generated in the FM3 melanoma cell line upon transfection of modified PX458 (Figure 3C and Figure S2A). Sanger sequencing (data not shown) and PCR amplification (Figure S2L) demonstrated the successful deletion of the miR-155-5p binding site (R3R9) resulting in an upregulation of tpn mRNA and protein levels when compared to the mock vector (PX458) (Figure 3D–F). As shown in Figure 3G, the tpn expression (green) was higher in the miR-155 group than in the other two groups, while the expression of H3K27me3 (red) was inversely correlated due to direct or indirect binding of tpn to H3K27me3 as shown by immunoprecipitation (Figure S2M), which strengthen the evidence that the binding site is part of a silencer.
Finally, to exclude that the miRNA-mediated activation of tpn was caused by RBPs, the four proteins HNRNPL, HNRNPC, IGF2BP1 and IGF2BP3, known to bind to the tpn 3′UTR, were explored. Compared to HNRNPC and HNRNPL, the IGF2BP1 and IGF2BP3 binding sites overlap with that of miR-155-5p (Figure S3A–D). Interestingly, the miR-155-5p binding site is a part of the IGF2BP1 and IGF2BP3 potential binding sites containing a GC- and an AU-rich element (Figure S3E). Concentration gradient silencing assays revealed an interaction of miR-155-5p and these RBPs (Figure S3F–H).
This study proposes a new unconventional function of miRNAs that enhances target transcription through binding a silencer thereby activating the downstream pathway(s) (Figure 4). Using miR-155-5p as a model, this is the first report (i) identifying a silencer in the tpn 3′UTR, which (ii) directly interacts with a non-coding RNA and (iii) has clinical relevance. These data extend miRNAs’ functions and add new insights to our knowledge of silencers.
Conceptualization: Yuan Wang, Barbara Seliger, Theresa Kordaß and Stefan Eichmüller. Methodology: Yuan Wang, Maria-Filothei Lazaridou, Chiara Massa, Theresa Kordaß, Christoforos K. Vaxevanis. Investigation: Yuan Wang, Maria-Filothei Lazaridou and Chiara Massa. Visualization: Yuan Wang, Maria-Filothei Lazaridou and Chiara Massa. Funding acquisition: Barbara Seliger and Stefan Eichmüller; Writing—original draft: Yuan Wang and Barbara Seliger; Writing—review & editing: Yuan Wang, Barbara Seliger, Theresa Kordaß, Stefan Eichmüller, Maria-Filothei Lazaridou, Chiara Massa, Christoforos K. Vaxevanis.
The authors declare no conflict of interest.
The work was funded by grants of the Deutsche Krebshilfe, grant 341025929 (Barbara Seliger) and the Deutsche Forschungsgemeinschaft, grant number: SE581/33-1.
期刊介绍:
Clinical and Translational Medicine (CTM) is an international, peer-reviewed, open-access journal dedicated to accelerating the translation of preclinical research into clinical applications and fostering communication between basic and clinical scientists. It highlights the clinical potential and application of various fields including biotechnologies, biomaterials, bioengineering, biomarkers, molecular medicine, omics science, bioinformatics, immunology, molecular imaging, drug discovery, regulation, and health policy. With a focus on the bench-to-bedside approach, CTM prioritizes studies and clinical observations that generate hypotheses relevant to patients and diseases, guiding investigations in cellular and molecular medicine. The journal encourages submissions from clinicians, researchers, policymakers, and industry professionals.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


