Joseph A. Daccache, Francis Eng, Lei Cao, Ning Ma, Stephen C. Ward, Thomas Schiano, Mark Miller, Daniel Herron, Anthony V. Azzara, Steven S. Pullen, Paolo Guarnieri, Costica Aloman, Andrea D. Branch
{"title":"单细胞 RNA 测序数据确定了可能在人体重要器官中无处不在的表达金属硫蛋白的巨噬细胞保守群体","authors":"Joseph A. Daccache, Francis Eng, Lei Cao, Ning Ma, Stephen C. Ward, Thomas Schiano, Mark Miller, Daniel Herron, Anthony V. Azzara, Steven S. Pullen, Paolo Guarnieri, Costica Aloman, Andrea D. Branch","doi":"10.1002/ctd2.348","DOIUrl":null,"url":null,"abstract":"<p>Dear Editor,</p><p>We used publicly available single-cell RNA sequencing data to identify conserved tissue macrophage populations in human organs. Among the subsets, we found a rare population of metallothionein-expressing macrophages that are present in all vital organs analyzed. We deeply phenotype this subset and determine their localization in the human liver.</p><p>In the first phase, we collected data from 10 livers,<span><sup>1</sup></span> 21 kidneys<span><sup>2</sup></span> and 60 lungs,<span><sup>3</sup></span> and selected myeloid-lineage cells using published annotations and lineage markers: CD68, S100, HLA-II, LYZ, C1Q, and CD74 (Figure S1). After removing dendritic cells, we subclustered monocytes and macrophages (liver, 8,197; kidney, 5,005; lung, 121,536), and identified eight subclusters in the monocyte-macrophage (mono-mac) space, defined by their differentially expressed genes (DEGs) (Figure 1A and Figure S2). We evaluated the relatedness of subclusters across organs by determining the Pearson correlations of fold-changes of 1126 DEGs that were significant [<i>p</i>.adj < .05 (Bonferroni)] in at least two organs (Figure 1B). Cross-organ correlations confirmed the relatedness of most subsets and were highest for monocytes and cycling macrophages.</p><p>A survey of the mono-mac subtypes present in at least two human organs revealed that the abundances of most were similar across organs, but MAC_RES was disproportionately large in the lung (alveolar macrophages) and absent from the kidney (Figure 1C). Subset abundances were compared between fibrotic and control organs and differed significantly for three: monocytes, MAC_RES and <i>SPP1</i><sup>+</sup> macrophages (Figure S2). While most of the eight subsets are widely known, we found an unexpected, small population of macrophages (∼2.5% of each organ) that highly expressed genes coding for metallothioneins (MAC_MT). Metallothioneins are low molecular weight, cysteine-rich proteins that bind metal ions. They enhance angiogenesis, protect against oxidative stress and metal toxicity, and modulate macrophage function.<span><sup>4</sup></span></p><p>Gene expression patterns of MAC_MTs were highly correlated across all three organs. Using a Wilcoxon rank-sum test to calculate DEGs, nine genes were significant [FC > 0, <i>p</i>.adj < .05 (Bonferroni)] in MAC_MTs in all three organs (Figure 1D). Genes encoding metallothioneins were highly expressed by MAC_MTs only, unlike <i>CTSL</i> and <i>SGK1</i>, which were expressed by multiple sub-clusters. Of interest, <i>MTF1</i> and <i>MTF2</i>, which encode metallothionein transcription factors, were minimally expressed in MAC_MTs but were highly expressed in the CYCLING subset, which may provide a local source of MAC_MTs.</p><p>In each organ, two gene signatures were developed, one distinguished MAC_MTs from other non-dendritic myeloid-lineage cells, and the other distinguished MAC_MTs from all other cells in the organ. To generate each signature, we selected DEGs [FC > 0.75, <i>p</i>.adj < .05 (Bonferroni)] whose expression was significantly greater in MAC_MTs than in the second-highest subset [FC > 0, <i>p</i>.adj < .05 (Bonferroni)]. This method generates gene signatures that can identify a subset of interest against a background of other cells. It removes genes that are highly expressed in other cell types and goes beyond an arbitrary number of top DEGs.</p><p>We next scored all individual cells based on their expression of both MAC_MT organ-specific gene signatures and compared scores across the subsets (Figure 1E and Table S1). The signatures of MAC_MTs vs. myeloid-lineage cells were dominated by <i>MT1</i> genes and <i>MT2A</i> but also included other immune response and metabolism-associated genes. The signature of liver MAC_MTs included <i>HAMP</i> (encodes hepcidin), an iron regulator. The signatures distinguished MAC_MTs from other myeloid-lineage cells, as demonstrated by UMAP overlays (Figure S3). The whole-organ signatures distinguished MAC_MTs of the liver and lung from all other cell types in the source organ (Figure S4), but did not distinguish kidney MAC_MTs with confidence, likely due to the high expression of metallothionein-encoding genes in kidney epithelial cells (Figure S5).</p><p>To infer the differentiation status of MAC_MTs, we performed pseudotemporal analysis with monocytes as the starting node. The trajectories suggest that MAC_MTs originate from monocytes and are not terminally differentiated (Figure 1F). Lineage tracing studies are needed to further define their origin and progeny.</p><p>To predict the potential functional roles of MAC_MTs in tissue, we inferred ligand-receptor interactions between MAC_MTs and all other cell types, focusing on ligands secreted by MAC_MTs. We identified 15 interactions conserved across tissues (Figure S6). Gene set enrichment analysis highlighted biological processes involving cellular migration, vascularization and angiogenesis. The genes driving these enrichments (<i>GRN</i>, <i>TNFSF12</i>, <i>IL1B</i>, <i>VEGFB</i> and <i>VEGFA</i>) may indicate that MAC_MTs participate in blood vessel formation and tissue remodelling (Figure 1G).</p><p>To gain a wider view of human MAC_MTs, we analyzed an additional dataset<span><sup>5</sup></span> in which the Macrophage-11 sub-cluster represents MAC_MTs (Figure 2A,B). Metallothionein gene expression was specific to this sub-cluster and present in all 10 vital organs (stomach, kidney, pancreas, colon, lung, ascites, tonsil, liver, breast, and skin) but absent from spleen, blood, and ascites fluid. MAC_MTs were a small percentage of mono-macs, less than 5% in all organs tested (Figure 2C). Metallothionein genes were highly expressed and specific to the Macrophage-11 cluster, as indicated by the high score of our MAC_MT myeloid signature (Figure 2D and Figure S7). These findings indicate that MAC_MTs are a distinctive subset present in multiple vital organs. They were previously reported to be limited to prostate, but have also been localized in lung.<span><sup>6, 7</sup></span></p><p>We used an advanced immunohistochemistry technique,<span><sup>8</sup></span> consecutive immunohistochemically staining on a single section, to localize MAC_MTs in human liver tissue (Figure 2E). The liver tissues of three patients with advanced liver disease were consecutively stained with an anti-CD68 antibody and an antibody that recognizes MT1 and MT2. Rare CD68+/MT+ cells were identified in all patients and were localized to the regions of ductular reactions that surround regenerative nodules in the cirrhotic liver and support neovascular development. These findings combine with the receptor-ligand data to suggest a role of MAC_MTs in neoangiogenesis.</p><p>In conclusion, using publicly available scRNA-sequencing data and immunohistochemistry, we transcriptionally profiled MAC_MTs, delineated shared marker genes of MAC_MTs, established their presence in transcriptomic data from multiple vital organs, identified predictors of MAC_MT function, and determined their localization in human liver. Our research suggests that MAC_MTs occur in the majority of vital human organs. Trajectory analysis suggests they differentiate locally within their resident organ, consistent with evidence that corneal tissue upregulates <i>MT</i> genes when cultured with monocyte-derived macrophages.<span><sup>9</sup></span> Our work sets the stage for mechanistic studies defining the functional role of MAC_MTs in tissue homeostasis and remodelling.</p><p><b>Joseph A. Daccache</b>: Conceptualization; investigation; visualization; interpretation and writing. <b>Francis Eng</b>: Conceptualization; investigation. <b>Lei Cao</b>: Methodology; validation. <b>Ning Ma, Stephen C. Ward, Thomas Schiano, Mark Miller and Daniel Herron</b>: Interpretation; resources. <b>Anthony V. Azzara, Steven S. Pullen and Paolo Guarnieri</b>: Conceptualization; Costica Aloman: Conceptualization, interpretation and funding. <b>Andrea D. Branch</b>: Conceptualization; interpretation; writing and funding acquisition.</p><p>Stephen C. Ward receives grant salary support from Boehringer Ingelheim, Ltd. Daniel Herron has received an honorarium from Intuitive in the past. Anthony V. Azzara and Steven S. Pullen are employees at Boehringer Ingelheim Pharmaceutical Inc. Paolo Guarnieri has stock ownership in 23 & Me. Andrea D. Branch advises the Center for Disease Analysis Foundation and the Icahn School of Medicine at Mount Sinai receives funding from Gilead and Pfizer to support laboratory research. Joseph A. Daccache, Francis Eng, Lei Cao, Ning Ma, Thomas Schiano, Mark Miller and Costica Aloman declare no conflict of interest.</p><p>This work was funded by Boehringer Ingelheim, NIOSH grants U010H012622 and U010H012263, NIAAA grants R01AA024762 and NCI grant 1U0CA288425, and the Prevent Cancer Foundation.</p><p>This study utilizes published datasets that have been deidentified and uploaded to the GEO and Zenodo archives. Human samples for IHC staining were collected under GCO numbers 15−1671 and 18−1512.</p>","PeriodicalId":72605,"journal":{"name":"Clinical and translational discovery","volume":"4 5","pages":""},"PeriodicalIF":1.9000,"publicationDate":"2024-08-27","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1002/ctd2.348","citationCount":"0","resultStr":"{\"title\":\"Single-cell RNA sequencing data identify a conserved population of metallothionein-expressing macrophages that may be ubiquitous in vital human organs\",\"authors\":\"Joseph A. Daccache, Francis Eng, Lei Cao, Ning Ma, Stephen C. Ward, Thomas Schiano, Mark Miller, Daniel Herron, Anthony V. Azzara, Steven S. Pullen, Paolo Guarnieri, Costica Aloman, Andrea D. Branch\",\"doi\":\"10.1002/ctd2.348\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p>Dear Editor,</p><p>We used publicly available single-cell RNA sequencing data to identify conserved tissue macrophage populations in human organs. Among the subsets, we found a rare population of metallothionein-expressing macrophages that are present in all vital organs analyzed. We deeply phenotype this subset and determine their localization in the human liver.</p><p>In the first phase, we collected data from 10 livers,<span><sup>1</sup></span> 21 kidneys<span><sup>2</sup></span> and 60 lungs,<span><sup>3</sup></span> and selected myeloid-lineage cells using published annotations and lineage markers: CD68, S100, HLA-II, LYZ, C1Q, and CD74 (Figure S1). After removing dendritic cells, we subclustered monocytes and macrophages (liver, 8,197; kidney, 5,005; lung, 121,536), and identified eight subclusters in the monocyte-macrophage (mono-mac) space, defined by their differentially expressed genes (DEGs) (Figure 1A and Figure S2). We evaluated the relatedness of subclusters across organs by determining the Pearson correlations of fold-changes of 1126 DEGs that were significant [<i>p</i>.adj < .05 (Bonferroni)] in at least two organs (Figure 1B). Cross-organ correlations confirmed the relatedness of most subsets and were highest for monocytes and cycling macrophages.</p><p>A survey of the mono-mac subtypes present in at least two human organs revealed that the abundances of most were similar across organs, but MAC_RES was disproportionately large in the lung (alveolar macrophages) and absent from the kidney (Figure 1C). Subset abundances were compared between fibrotic and control organs and differed significantly for three: monocytes, MAC_RES and <i>SPP1</i><sup>+</sup> macrophages (Figure S2). While most of the eight subsets are widely known, we found an unexpected, small population of macrophages (∼2.5% of each organ) that highly expressed genes coding for metallothioneins (MAC_MT). Metallothioneins are low molecular weight, cysteine-rich proteins that bind metal ions. They enhance angiogenesis, protect against oxidative stress and metal toxicity, and modulate macrophage function.<span><sup>4</sup></span></p><p>Gene expression patterns of MAC_MTs were highly correlated across all three organs. Using a Wilcoxon rank-sum test to calculate DEGs, nine genes were significant [FC > 0, <i>p</i>.adj < .05 (Bonferroni)] in MAC_MTs in all three organs (Figure 1D). Genes encoding metallothioneins were highly expressed by MAC_MTs only, unlike <i>CTSL</i> and <i>SGK1</i>, which were expressed by multiple sub-clusters. Of interest, <i>MTF1</i> and <i>MTF2</i>, which encode metallothionein transcription factors, were minimally expressed in MAC_MTs but were highly expressed in the CYCLING subset, which may provide a local source of MAC_MTs.</p><p>In each organ, two gene signatures were developed, one distinguished MAC_MTs from other non-dendritic myeloid-lineage cells, and the other distinguished MAC_MTs from all other cells in the organ. To generate each signature, we selected DEGs [FC > 0.75, <i>p</i>.adj < .05 (Bonferroni)] whose expression was significantly greater in MAC_MTs than in the second-highest subset [FC > 0, <i>p</i>.adj < .05 (Bonferroni)]. This method generates gene signatures that can identify a subset of interest against a background of other cells. It removes genes that are highly expressed in other cell types and goes beyond an arbitrary number of top DEGs.</p><p>We next scored all individual cells based on their expression of both MAC_MT organ-specific gene signatures and compared scores across the subsets (Figure 1E and Table S1). The signatures of MAC_MTs vs. myeloid-lineage cells were dominated by <i>MT1</i> genes and <i>MT2A</i> but also included other immune response and metabolism-associated genes. The signature of liver MAC_MTs included <i>HAMP</i> (encodes hepcidin), an iron regulator. The signatures distinguished MAC_MTs from other myeloid-lineage cells, as demonstrated by UMAP overlays (Figure S3). The whole-organ signatures distinguished MAC_MTs of the liver and lung from all other cell types in the source organ (Figure S4), but did not distinguish kidney MAC_MTs with confidence, likely due to the high expression of metallothionein-encoding genes in kidney epithelial cells (Figure S5).</p><p>To infer the differentiation status of MAC_MTs, we performed pseudotemporal analysis with monocytes as the starting node. The trajectories suggest that MAC_MTs originate from monocytes and are not terminally differentiated (Figure 1F). Lineage tracing studies are needed to further define their origin and progeny.</p><p>To predict the potential functional roles of MAC_MTs in tissue, we inferred ligand-receptor interactions between MAC_MTs and all other cell types, focusing on ligands secreted by MAC_MTs. We identified 15 interactions conserved across tissues (Figure S6). Gene set enrichment analysis highlighted biological processes involving cellular migration, vascularization and angiogenesis. The genes driving these enrichments (<i>GRN</i>, <i>TNFSF12</i>, <i>IL1B</i>, <i>VEGFB</i> and <i>VEGFA</i>) may indicate that MAC_MTs participate in blood vessel formation and tissue remodelling (Figure 1G).</p><p>To gain a wider view of human MAC_MTs, we analyzed an additional dataset<span><sup>5</sup></span> in which the Macrophage-11 sub-cluster represents MAC_MTs (Figure 2A,B). Metallothionein gene expression was specific to this sub-cluster and present in all 10 vital organs (stomach, kidney, pancreas, colon, lung, ascites, tonsil, liver, breast, and skin) but absent from spleen, blood, and ascites fluid. MAC_MTs were a small percentage of mono-macs, less than 5% in all organs tested (Figure 2C). Metallothionein genes were highly expressed and specific to the Macrophage-11 cluster, as indicated by the high score of our MAC_MT myeloid signature (Figure 2D and Figure S7). These findings indicate that MAC_MTs are a distinctive subset present in multiple vital organs. They were previously reported to be limited to prostate, but have also been localized in lung.<span><sup>6, 7</sup></span></p><p>We used an advanced immunohistochemistry technique,<span><sup>8</sup></span> consecutive immunohistochemically staining on a single section, to localize MAC_MTs in human liver tissue (Figure 2E). The liver tissues of three patients with advanced liver disease were consecutively stained with an anti-CD68 antibody and an antibody that recognizes MT1 and MT2. Rare CD68+/MT+ cells were identified in all patients and were localized to the regions of ductular reactions that surround regenerative nodules in the cirrhotic liver and support neovascular development. These findings combine with the receptor-ligand data to suggest a role of MAC_MTs in neoangiogenesis.</p><p>In conclusion, using publicly available scRNA-sequencing data and immunohistochemistry, we transcriptionally profiled MAC_MTs, delineated shared marker genes of MAC_MTs, established their presence in transcriptomic data from multiple vital organs, identified predictors of MAC_MT function, and determined their localization in human liver. Our research suggests that MAC_MTs occur in the majority of vital human organs. Trajectory analysis suggests they differentiate locally within their resident organ, consistent with evidence that corneal tissue upregulates <i>MT</i> genes when cultured with monocyte-derived macrophages.<span><sup>9</sup></span> Our work sets the stage for mechanistic studies defining the functional role of MAC_MTs in tissue homeostasis and remodelling.</p><p><b>Joseph A. Daccache</b>: Conceptualization; investigation; visualization; interpretation and writing. <b>Francis Eng</b>: Conceptualization; investigation. <b>Lei Cao</b>: Methodology; validation. <b>Ning Ma, Stephen C. Ward, Thomas Schiano, Mark Miller and Daniel Herron</b>: Interpretation; resources. <b>Anthony V. Azzara, Steven S. Pullen and Paolo Guarnieri</b>: Conceptualization; Costica Aloman: Conceptualization, interpretation and funding. <b>Andrea D. Branch</b>: Conceptualization; interpretation; writing and funding acquisition.</p><p>Stephen C. Ward receives grant salary support from Boehringer Ingelheim, Ltd. Daniel Herron has received an honorarium from Intuitive in the past. Anthony V. Azzara and Steven S. Pullen are employees at Boehringer Ingelheim Pharmaceutical Inc. Paolo Guarnieri has stock ownership in 23 & Me. Andrea D. Branch advises the Center for Disease Analysis Foundation and the Icahn School of Medicine at Mount Sinai receives funding from Gilead and Pfizer to support laboratory research. Joseph A. Daccache, Francis Eng, Lei Cao, Ning Ma, Thomas Schiano, Mark Miller and Costica Aloman declare no conflict of interest.</p><p>This work was funded by Boehringer Ingelheim, NIOSH grants U010H012622 and U010H012263, NIAAA grants R01AA024762 and NCI grant 1U0CA288425, and the Prevent Cancer Foundation.</p><p>This study utilizes published datasets that have been deidentified and uploaded to the GEO and Zenodo archives. Human samples for IHC staining were collected under GCO numbers 15−1671 and 18−1512.</p>\",\"PeriodicalId\":72605,\"journal\":{\"name\":\"Clinical and translational discovery\",\"volume\":\"4 5\",\"pages\":\"\"},\"PeriodicalIF\":1.9000,\"publicationDate\":\"2024-08-27\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://onlinelibrary.wiley.com/doi/epdf/10.1002/ctd2.348\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"Clinical and translational discovery\",\"FirstCategoryId\":\"1085\",\"ListUrlMain\":\"https://onlinelibrary.wiley.com/doi/10.1002/ctd2.348\",\"RegionNum\":0,\"RegionCategory\":null,\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"\",\"JCRName\":\"\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"Clinical and translational discovery","FirstCategoryId":"1085","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1002/ctd2.348","RegionNum":0,"RegionCategory":null,"ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"","JCRName":"","Score":null,"Total":0}
Single-cell RNA sequencing data identify a conserved population of metallothionein-expressing macrophages that may be ubiquitous in vital human organs
Dear Editor,
We used publicly available single-cell RNA sequencing data to identify conserved tissue macrophage populations in human organs. Among the subsets, we found a rare population of metallothionein-expressing macrophages that are present in all vital organs analyzed. We deeply phenotype this subset and determine their localization in the human liver.
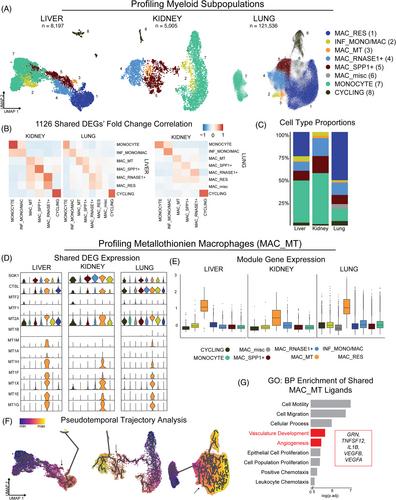
In the first phase, we collected data from 10 livers,1 21 kidneys2 and 60 lungs,3 and selected myeloid-lineage cells using published annotations and lineage markers: CD68, S100, HLA-II, LYZ, C1Q, and CD74 (Figure S1). After removing dendritic cells, we subclustered monocytes and macrophages (liver, 8,197; kidney, 5,005; lung, 121,536), and identified eight subclusters in the monocyte-macrophage (mono-mac) space, defined by their differentially expressed genes (DEGs) (Figure 1A and Figure S2). We evaluated the relatedness of subclusters across organs by determining the Pearson correlations of fold-changes of 1126 DEGs that were significant [p.adj < .05 (Bonferroni)] in at least two organs (Figure 1B). Cross-organ correlations confirmed the relatedness of most subsets and were highest for monocytes and cycling macrophages.
A survey of the mono-mac subtypes present in at least two human organs revealed that the abundances of most were similar across organs, but MAC_RES was disproportionately large in the lung (alveolar macrophages) and absent from the kidney (Figure 1C). Subset abundances were compared between fibrotic and control organs and differed significantly for three: monocytes, MAC_RES and SPP1+ macrophages (Figure S2). While most of the eight subsets are widely known, we found an unexpected, small population of macrophages (∼2.5% of each organ) that highly expressed genes coding for metallothioneins (MAC_MT). Metallothioneins are low molecular weight, cysteine-rich proteins that bind metal ions. They enhance angiogenesis, protect against oxidative stress and metal toxicity, and modulate macrophage function.4
Gene expression patterns of MAC_MTs were highly correlated across all three organs. Using a Wilcoxon rank-sum test to calculate DEGs, nine genes were significant [FC > 0, p.adj < .05 (Bonferroni)] in MAC_MTs in all three organs (Figure 1D). Genes encoding metallothioneins were highly expressed by MAC_MTs only, unlike CTSL and SGK1, which were expressed by multiple sub-clusters. Of interest, MTF1 and MTF2, which encode metallothionein transcription factors, were minimally expressed in MAC_MTs but were highly expressed in the CYCLING subset, which may provide a local source of MAC_MTs.
In each organ, two gene signatures were developed, one distinguished MAC_MTs from other non-dendritic myeloid-lineage cells, and the other distinguished MAC_MTs from all other cells in the organ. To generate each signature, we selected DEGs [FC > 0.75, p.adj < .05 (Bonferroni)] whose expression was significantly greater in MAC_MTs than in the second-highest subset [FC > 0, p.adj < .05 (Bonferroni)]. This method generates gene signatures that can identify a subset of interest against a background of other cells. It removes genes that are highly expressed in other cell types and goes beyond an arbitrary number of top DEGs.
We next scored all individual cells based on their expression of both MAC_MT organ-specific gene signatures and compared scores across the subsets (Figure 1E and Table S1). The signatures of MAC_MTs vs. myeloid-lineage cells were dominated by MT1 genes and MT2A but also included other immune response and metabolism-associated genes. The signature of liver MAC_MTs included HAMP (encodes hepcidin), an iron regulator. The signatures distinguished MAC_MTs from other myeloid-lineage cells, as demonstrated by UMAP overlays (Figure S3). The whole-organ signatures distinguished MAC_MTs of the liver and lung from all other cell types in the source organ (Figure S4), but did not distinguish kidney MAC_MTs with confidence, likely due to the high expression of metallothionein-encoding genes in kidney epithelial cells (Figure S5).
To infer the differentiation status of MAC_MTs, we performed pseudotemporal analysis with monocytes as the starting node. The trajectories suggest that MAC_MTs originate from monocytes and are not terminally differentiated (Figure 1F). Lineage tracing studies are needed to further define their origin and progeny.
To predict the potential functional roles of MAC_MTs in tissue, we inferred ligand-receptor interactions between MAC_MTs and all other cell types, focusing on ligands secreted by MAC_MTs. We identified 15 interactions conserved across tissues (Figure S6). Gene set enrichment analysis highlighted biological processes involving cellular migration, vascularization and angiogenesis. The genes driving these enrichments (GRN, TNFSF12, IL1B, VEGFB and VEGFA) may indicate that MAC_MTs participate in blood vessel formation and tissue remodelling (Figure 1G).
To gain a wider view of human MAC_MTs, we analyzed an additional dataset5 in which the Macrophage-11 sub-cluster represents MAC_MTs (Figure 2A,B). Metallothionein gene expression was specific to this sub-cluster and present in all 10 vital organs (stomach, kidney, pancreas, colon, lung, ascites, tonsil, liver, breast, and skin) but absent from spleen, blood, and ascites fluid. MAC_MTs were a small percentage of mono-macs, less than 5% in all organs tested (Figure 2C). Metallothionein genes were highly expressed and specific to the Macrophage-11 cluster, as indicated by the high score of our MAC_MT myeloid signature (Figure 2D and Figure S7). These findings indicate that MAC_MTs are a distinctive subset present in multiple vital organs. They were previously reported to be limited to prostate, but have also been localized in lung.6, 7
We used an advanced immunohistochemistry technique,8 consecutive immunohistochemically staining on a single section, to localize MAC_MTs in human liver tissue (Figure 2E). The liver tissues of three patients with advanced liver disease were consecutively stained with an anti-CD68 antibody and an antibody that recognizes MT1 and MT2. Rare CD68+/MT+ cells were identified in all patients and were localized to the regions of ductular reactions that surround regenerative nodules in the cirrhotic liver and support neovascular development. These findings combine with the receptor-ligand data to suggest a role of MAC_MTs in neoangiogenesis.
In conclusion, using publicly available scRNA-sequencing data and immunohistochemistry, we transcriptionally profiled MAC_MTs, delineated shared marker genes of MAC_MTs, established their presence in transcriptomic data from multiple vital organs, identified predictors of MAC_MT function, and determined their localization in human liver. Our research suggests that MAC_MTs occur in the majority of vital human organs. Trajectory analysis suggests they differentiate locally within their resident organ, consistent with evidence that corneal tissue upregulates MT genes when cultured with monocyte-derived macrophages.9 Our work sets the stage for mechanistic studies defining the functional role of MAC_MTs in tissue homeostasis and remodelling.
Joseph A. Daccache: Conceptualization; investigation; visualization; interpretation and writing. Francis Eng: Conceptualization; investigation. Lei Cao: Methodology; validation. Ning Ma, Stephen C. Ward, Thomas Schiano, Mark Miller and Daniel Herron: Interpretation; resources. Anthony V. Azzara, Steven S. Pullen and Paolo Guarnieri: Conceptualization; Costica Aloman: Conceptualization, interpretation and funding. Andrea D. Branch: Conceptualization; interpretation; writing and funding acquisition.
Stephen C. Ward receives grant salary support from Boehringer Ingelheim, Ltd. Daniel Herron has received an honorarium from Intuitive in the past. Anthony V. Azzara and Steven S. Pullen are employees at Boehringer Ingelheim Pharmaceutical Inc. Paolo Guarnieri has stock ownership in 23 & Me. Andrea D. Branch advises the Center for Disease Analysis Foundation and the Icahn School of Medicine at Mount Sinai receives funding from Gilead and Pfizer to support laboratory research. Joseph A. Daccache, Francis Eng, Lei Cao, Ning Ma, Thomas Schiano, Mark Miller and Costica Aloman declare no conflict of interest.
This work was funded by Boehringer Ingelheim, NIOSH grants U010H012622 and U010H012263, NIAAA grants R01AA024762 and NCI grant 1U0CA288425, and the Prevent Cancer Foundation.
This study utilizes published datasets that have been deidentified and uploaded to the GEO and Zenodo archives. Human samples for IHC staining were collected under GCO numbers 15−1671 and 18−1512.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


