{"title":"枯草芽孢杆菌脂肪酸生物合成启动酶的晶体结构","authors":"Christopher D. Radka , Charles O. Rock","doi":"10.1016/j.jsb.2024.108065","DOIUrl":null,"url":null,"abstract":"<div><p>Bacteria use the fatty acid composition of membrane lipids to maintain homeostasis of the bilayer. β-Ketoacyl-ACP synthase III (FabH) initiates fatty acid biosynthesis and is the primary determinant of the fatty acid composition. FabH condenses malonyl-acyl carrier protein with an acyl-Coenzyme A primer to form β -ketoacyl-acyl carrier protein which is used to make substrates for lipid synthesis. The acyl-Coenzyme A primer determines whether an acyl chain in the membrane has iso, anteiso, or no branching (straight chain) and biophysical properties of the membrane. The soil bacterium <em>Bacillus subtilis</em> encodes two copies of FabH (<em>Bs</em>FabHA and <em>Bs</em>FabHB), and here we solve their crystal structures. The substrate-free 1.85 Å and 2.40 Å structures of <em>Bs</em>FabHA and <em>Bs</em>FabHB show both enzymes have similar residues that line the active site but differ in the architecture surrounding the catalytic residues and oxyanion hole. Branching in the <em>Bs</em>FabHB active site may better accommodate the structure of an iso-branched acyl-Coenzyme A molecule and thus confer superior utilization to <em>Bs</em>FabHA for this primer type. The 2.02 Å structure of <em>Bs</em>FabHA•Coenzyme A shows how the active site architecture changes after binding the first substrate. The other notable difference is an amino acid insertion in <em>Bs</em>FabHB that extends a cap that covers the dimer interface. The cap topology is diverse across FabH structures and appears to be a distinguishing feature. FabH enzymes have variable sensitivity to natural product inhibitors and the availability of crystal structures help clarify how nature designs antimicrobials that differentially target FabH homologs.</p></div>","PeriodicalId":17074,"journal":{"name":"Journal of structural biology","volume":"216 1","pages":"Article 108065"},"PeriodicalIF":3.0000,"publicationDate":"2024-02-03","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://www.sciencedirect.com/science/article/pii/S1047847724000054/pdfft?md5=241a6f10088e07721d7498722a671e26&pid=1-s2.0-S1047847724000054-main.pdf","citationCount":"0","resultStr":"{\"title\":\"Crystal structures of the fatty acid biosynthesis initiation enzymes in Bacillus subtilis\",\"authors\":\"Christopher D. Radka , Charles O. Rock\",\"doi\":\"10.1016/j.jsb.2024.108065\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<div><p>Bacteria use the fatty acid composition of membrane lipids to maintain homeostasis of the bilayer. β-Ketoacyl-ACP synthase III (FabH) initiates fatty acid biosynthesis and is the primary determinant of the fatty acid composition. FabH condenses malonyl-acyl carrier protein with an acyl-Coenzyme A primer to form β -ketoacyl-acyl carrier protein which is used to make substrates for lipid synthesis. The acyl-Coenzyme A primer determines whether an acyl chain in the membrane has iso, anteiso, or no branching (straight chain) and biophysical properties of the membrane. The soil bacterium <em>Bacillus subtilis</em> encodes two copies of FabH (<em>Bs</em>FabHA and <em>Bs</em>FabHB), and here we solve their crystal structures. The substrate-free 1.85 Å and 2.40 Å structures of <em>Bs</em>FabHA and <em>Bs</em>FabHB show both enzymes have similar residues that line the active site but differ in the architecture surrounding the catalytic residues and oxyanion hole. Branching in the <em>Bs</em>FabHB active site may better accommodate the structure of an iso-branched acyl-Coenzyme A molecule and thus confer superior utilization to <em>Bs</em>FabHA for this primer type. The 2.02 Å structure of <em>Bs</em>FabHA•Coenzyme A shows how the active site architecture changes after binding the first substrate. The other notable difference is an amino acid insertion in <em>Bs</em>FabHB that extends a cap that covers the dimer interface. The cap topology is diverse across FabH structures and appears to be a distinguishing feature. FabH enzymes have variable sensitivity to natural product inhibitors and the availability of crystal structures help clarify how nature designs antimicrobials that differentially target FabH homologs.</p></div>\",\"PeriodicalId\":17074,\"journal\":{\"name\":\"Journal of structural biology\",\"volume\":\"216 1\",\"pages\":\"Article 108065\"},\"PeriodicalIF\":3.0000,\"publicationDate\":\"2024-02-03\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://www.sciencedirect.com/science/article/pii/S1047847724000054/pdfft?md5=241a6f10088e07721d7498722a671e26&pid=1-s2.0-S1047847724000054-main.pdf\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"Journal of structural biology\",\"FirstCategoryId\":\"99\",\"ListUrlMain\":\"https://www.sciencedirect.com/science/article/pii/S1047847724000054\",\"RegionNum\":3,\"RegionCategory\":\"生物学\",\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"Q3\",\"JCRName\":\"BIOCHEMISTRY & MOLECULAR BIOLOGY\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"Journal of structural biology","FirstCategoryId":"99","ListUrlMain":"https://www.sciencedirect.com/science/article/pii/S1047847724000054","RegionNum":3,"RegionCategory":"生物学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q3","JCRName":"BIOCHEMISTRY & MOLECULAR BIOLOGY","Score":null,"Total":0}
引用次数: 0
摘要
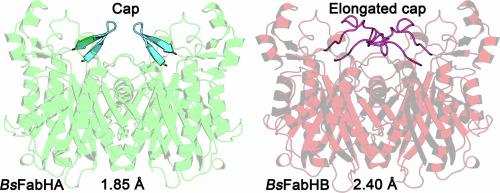
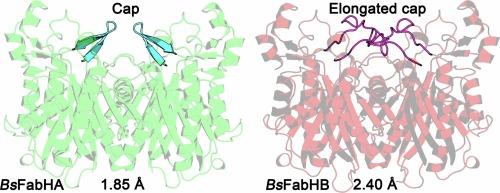
细菌利用膜脂的脂肪酸组成来维持双分子层的平衡。β-酮酰-ACP 合酶 III(FabH)启动脂肪酸的生物合成,是脂肪酸组成的主要决定因素。FabH将丙二酰-酰基载体蛋白与酰基辅酶A引物缩合,形成β-酮酰-酰基载体蛋白,用于制造脂质合成的底物。酰基辅酶 A 引物决定了膜中的酰基链是异链、前异链还是无分支(直链)以及膜的生物物理特性。土壤杆菌编码了两份 FabH(BsFabHA 和 BsFabHB),我们在此解析了它们的晶体结构。BsFabHA 和 BsFabHB 的无底物 1.85 Å 和 2.40 Å 晶体结构显示,两种酶的活性位点残基相似,但催化残基和氧阴离子孔周围的结构不同。BsFabHB 活性位点的分支可能更适合前异分支酰基辅酶 A 分子的结构,从而使 BsFabHA 更好地利用这种引物类型。BsFabHA-Coenzyme A 的 2.02 Å 结构显示了结合第一个底物后活性位点结构的变化。另一个显著的不同点是 BsFabHB 中的一个氨基酸插入物延伸出一个覆盖二聚体界面的帽。帽的拓扑结构在 FabH 结构中多种多样,似乎是一个显著特征。FabH酶对天然产物抑制剂的敏感性各不相同,晶体结构的可用性有助于阐明大自然是如何设计出针对不同FabH同源物的抗菌剂的。
Crystal structures of the fatty acid biosynthesis initiation enzymes in Bacillus subtilis
Bacteria use the fatty acid composition of membrane lipids to maintain homeostasis of the bilayer. β-Ketoacyl-ACP synthase III (FabH) initiates fatty acid biosynthesis and is the primary determinant of the fatty acid composition. FabH condenses malonyl-acyl carrier protein with an acyl-Coenzyme A primer to form β -ketoacyl-acyl carrier protein which is used to make substrates for lipid synthesis. The acyl-Coenzyme A primer determines whether an acyl chain in the membrane has iso, anteiso, or no branching (straight chain) and biophysical properties of the membrane. The soil bacterium Bacillus subtilis encodes two copies of FabH (BsFabHA and BsFabHB), and here we solve their crystal structures. The substrate-free 1.85 Å and 2.40 Å structures of BsFabHA and BsFabHB show both enzymes have similar residues that line the active site but differ in the architecture surrounding the catalytic residues and oxyanion hole. Branching in the BsFabHB active site may better accommodate the structure of an iso-branched acyl-Coenzyme A molecule and thus confer superior utilization to BsFabHA for this primer type. The 2.02 Å structure of BsFabHA•Coenzyme A shows how the active site architecture changes after binding the first substrate. The other notable difference is an amino acid insertion in BsFabHB that extends a cap that covers the dimer interface. The cap topology is diverse across FabH structures and appears to be a distinguishing feature. FabH enzymes have variable sensitivity to natural product inhibitors and the availability of crystal structures help clarify how nature designs antimicrobials that differentially target FabH homologs.
期刊介绍:
Journal of Structural Biology (JSB) has an open access mirror journal, the Journal of Structural Biology: X (JSBX), sharing the same aims and scope, editorial team, submission system and rigorous peer review. Since both journals share the same editorial system, you may submit your manuscript via either journal homepage. You will be prompted during submission (and revision) to choose in which to publish your article. The editors and reviewers are not aware of the choice you made until the article has been published online. JSB and JSBX publish papers dealing with the structural analysis of living material at every level of organization by all methods that lead to an understanding of biological function in terms of molecular and supermolecular structure.
Techniques covered include:
• Light microscopy including confocal microscopy
• All types of electron microscopy
• X-ray diffraction
• Nuclear magnetic resonance
• Scanning force microscopy, scanning probe microscopy, and tunneling microscopy
• Digital image processing
• Computational insights into structure

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


