{"title":"Structural and catalytic alteration of sarcosine oxidase through reconstruction with coenzyme-like ligands","authors":"Yu Xin, Mengling Zheng, Qing Wang, Liushen Lu, Ling Zhang, Yanjun Tong, Wu Wang","doi":"10.1016/j.molcatb.2017.01.011","DOIUrl":null,"url":null,"abstract":"<div><p>A sarcosine oxidase (SOX) gene from <em>Bacillus</em> sp. (AY626822.2) was expressed in <em>Escherichia coli</em> BL21 (DE3) in the form of inclusion bodies. A 3D model of SOX was then built and refined, and molecular docking was used to investigate the interactions between SOX and natural or coenzyme-like ligands, including flavin adenine dinucleotide (FAD); flavin mononucleotide (FMN); riboflavin; isoalloxazine; 7-methyl-8-chloro-10-(1′-<span>d</span>-ribityl) isoalloxazine (7-M-8-C); 7-bromo-8-methyl-10-(1′-<span>d</span>-ribityl) isoalloxazine (7-B-8-M); 7-methyl-8-bromo-10-(1′-<span>d</span>-ribityl) isoalloxazine (7-M-8-B); 7-chloro-8-ethyl-10-(1′-<span>d</span>-ribityl) isoalloxazine (7-C-8-E); 7,8-diethyl-10-(1′-<span>d</span>-ribityl) isoalloxazine (7,8-D); and 3-methyl-7,8-dimethyl-10-(1′-<span>d</span>-ribityl) isoalloxazine (3-M-7,8-D). Unfolded SOX was extracted from inclusion bodies, and reconstructed with these ligands <em>via</em><span> a refolding process. The reconstructed enzymes were then subjected to structural and catalytic analysis. After structural simulation, refinement, and molecular docking, all ligands were able to recognize the coenzyme site of SOX. In addition, when the position 7- or 8-site of the compounds was modified, new pi-cation/sigma interactions were formed in the SOX-ligand complex. Fluorescent detection revealed that all the ligands could be successfully reconstructed with unfolded SOX. Circular dichroism (CD) spectra and nano differential scanning calorimetry (DSC) analysis indicated that the loss of phosphoric acid and adeninein natural coenzymes could significantly reduce the α-helix content, transition temperature (T</span><sub>m</sub>), and calorimetric enthalpy (ΔH). In addition, although reconstruction with the position 7- or 8-site modified compounds led to variations in secondary structure, no significant shifts in T<sub>m</sub> and ΔH were observed. Furthermore, in the evaluation of catalytic kinetic parameters, when SOX was reconstructed with ligands containing halogen atoms at the 7- or 8-sites, much higher relative specificities in the presence of organic solvents were noted.</p></div>","PeriodicalId":16416,"journal":{"name":"Journal of Molecular Catalysis B-enzymatic","volume":"133 ","pages":"Pages S250-S258"},"PeriodicalIF":0.0000,"publicationDate":"2016-11-01","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://sci-hub-pdf.com/10.1016/j.molcatb.2017.01.011","citationCount":"8","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"Journal of Molecular Catalysis B-enzymatic","FirstCategoryId":"1085","ListUrlMain":"https://www.sciencedirect.com/science/article/pii/S1381117717300115","RegionNum":0,"RegionCategory":null,"ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q2","JCRName":"Chemical Engineering","Score":null,"Total":0}
引用次数: 8
Abstract
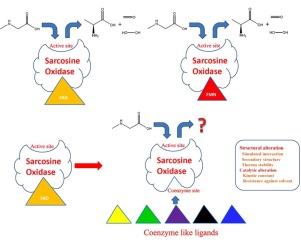
A sarcosine oxidase (SOX) gene from Bacillus sp. (AY626822.2) was expressed in Escherichia coli BL21 (DE3) in the form of inclusion bodies. A 3D model of SOX was then built and refined, and molecular docking was used to investigate the interactions between SOX and natural or coenzyme-like ligands, including flavin adenine dinucleotide (FAD); flavin mononucleotide (FMN); riboflavin; isoalloxazine; 7-methyl-8-chloro-10-(1′-d-ribityl) isoalloxazine (7-M-8-C); 7-bromo-8-methyl-10-(1′-d-ribityl) isoalloxazine (7-B-8-M); 7-methyl-8-bromo-10-(1′-d-ribityl) isoalloxazine (7-M-8-B); 7-chloro-8-ethyl-10-(1′-d-ribityl) isoalloxazine (7-C-8-E); 7,8-diethyl-10-(1′-d-ribityl) isoalloxazine (7,8-D); and 3-methyl-7,8-dimethyl-10-(1′-d-ribityl) isoalloxazine (3-M-7,8-D). Unfolded SOX was extracted from inclusion bodies, and reconstructed with these ligands via a refolding process. The reconstructed enzymes were then subjected to structural and catalytic analysis. After structural simulation, refinement, and molecular docking, all ligands were able to recognize the coenzyme site of SOX. In addition, when the position 7- or 8-site of the compounds was modified, new pi-cation/sigma interactions were formed in the SOX-ligand complex. Fluorescent detection revealed that all the ligands could be successfully reconstructed with unfolded SOX. Circular dichroism (CD) spectra and nano differential scanning calorimetry (DSC) analysis indicated that the loss of phosphoric acid and adeninein natural coenzymes could significantly reduce the α-helix content, transition temperature (Tm), and calorimetric enthalpy (ΔH). In addition, although reconstruction with the position 7- or 8-site modified compounds led to variations in secondary structure, no significant shifts in Tm and ΔH were observed. Furthermore, in the evaluation of catalytic kinetic parameters, when SOX was reconstructed with ligands containing halogen atoms at the 7- or 8-sites, much higher relative specificities in the presence of organic solvents were noted.
期刊介绍:
Journal of Molecular Catalysis B: Enzymatic is an international forum for researchers and product developers in the applications of whole-cell and cell-free enzymes as catalysts in organic synthesis. Emphasis is on mechanistic and synthetic aspects of the biocatalytic transformation.
Papers should report novel and significant advances in one or more of the following topics;
Applied and fundamental studies of enzymes used for biocatalysis;
Industrial applications of enzymatic processes, e.g. in fine chemical synthesis;
Chemo-, regio- and enantioselective transformations;
Screening for biocatalysts;
Integration of biocatalytic and chemical steps in organic syntheses;
Novel biocatalysts, e.g. enzymes from extremophiles and catalytic antibodies;
Enzyme immobilization and stabilization, particularly in non-conventional media;
Bioprocess engineering aspects, e.g. membrane bioreactors;
Improvement of catalytic performance of enzymes, e.g. by protein engineering or chemical modification;
Structural studies, including computer simulation, relating to substrate specificity and reaction selectivity;
Biomimetic studies related to enzymatic transformations.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


