{"title":"Understanding the interplay between N6-methyladenine RNA methylation and noncoding RNAs in kidney disease","authors":"Saiya Zhu, Dong Zhou","doi":"10.1002/ctd2.246","DOIUrl":null,"url":null,"abstract":"<p>First discovered in 1974,<span><sup>1</sup></span> N6-methyladenosine (m<sup>6</sup>A) is the most prevalent epigenetic modification in eukaryotic mRNAs at the posttranscriptional level under pathophysiological conditions. The m<sup>6</sup>A modification entails methylation of adenine at the nitrogen 6 (N6) position. Three classes of enzymes mediate the process of m<sup>6</sup>A modification, including RNA methyltransferases (writers), demethylases (erasers), and methylation recognition proteins (readers).<span><sup>2</sup></span> Writers can form a multicomponent m<sup>6</sup>A methyltransferase complex (MTC) that catalyzes RNA modification by transferring methyl groups to RNA. In MTC, METTL3, METTL14 and WTAP are core components. Serving as erasers, as the name implies, demethylases (FTO and ALKBH5) are responsible for removing S-adenosyl methionine from adenines in RNA. At the same time, readers (YTHDF, YTHDC and eIF3) can recognize m<sup>6</sup>A-modified RNAs and bind the m<sup>6</sup>A modification, subsequently determining the fate of the RNA. Therefore, m<sup>6</sup>A RNA modification is considered invertible and dynamic. In recent years, the advent of advanced high-throughput sequencing has facilitated the identification of m<sup>6</sup>A.</p><p>In general, m<sup>6</sup>A presents 0.4%−0.6% of all cellular RNAs, including mRNAs, noncoding RNAs (ncRNAs), and circular RNAs (circRNAs).<span><sup>3</sup></span> In a recent publication in <i>Clinical and Translational Medicine</i>, Ni et al. reported that METTL3 promoted the m<sup>6</sup>A modification of Ena/VASP-like (EVL) mRNA and subsequently activated the transforming growth factor-beta 1 (TGF-β1) signalling pathway, which ultimately aggravated kidney fibrosis.<span><sup>4</sup></span> Beyond regulating mRNA-coding genes, m<sup>6</sup>A modification exhibited remarkable capacity in modulating the function of ncRNAs to dictate kidney fate after various insults, especially in kidney carcinoma. Conversely, ncRNAs also participate in the regulation of m<sup>6</sup>A modification amid kidney disease progression. Therefore, fully understanding the crosstalk between m<sup>6</sup>A modification and ncRNAs will supplement a piece of the puzzle of the pathogenesis of non-tumour kidney diseases and shed light on developing effective therapeutic strategies for these medical catastrophes.</p><p>ncRNAs, mainly comprised of micro RNAs (miRNAs), long noncoding RNAs (lncRNAs), and circRNAs, account for a large class of transcripts in mammalian genomes without protein-coding potential. Despite this, they are crucial regulators of diverse physiological and pathological processes. The m<sup>6</sup>A modification of ncRNAs controls RNA stability, processing, and transport. As illustrated in Figure 1, accumulating studies have shown that m<sup>6</sup>A-modified ncRNAs cause kidney disorders through multiple dysregulated signalling pathways in diseased kidneys. Because the evidence of m<sup>6</sup>A modification of circRNAs remains lacking, we mainly focus on introducing the role of miRNAs and lncRNAs in the kidney system. miRNAs make up the majority of ncRNAs, and the role of m<sup>6</sup>A modification of miRNA is complex and diverse. For instance, in vitro, METTL3 positively modulated miR-873-5p to attenuate oxidative stress and apoptosis by regulating the Keap1/Nrf2 pathway in the cultured proximal tubular cells stimulated with colistin.<span><sup>5</sup></span> In vivo, METTL3-mediated m<sup>6</sup>A modification promoted miR-21-5p maturation. Matured miR-21-5p subsequently activated the SPRY1/ERK/NF-κB axis in the obstructive kidneys, exacerbating inflammation and ultimately driving kidney fibrosis.<span><sup>6</sup></span> As depicted in these findings, m<sup>6</sup>A modification may exert opposite regulatory roles at different stages of miRNA maturation. Therefore, obtaining a full image of m<sup>6</sup>A modification of miRNA is required to elucidate the regulatory mechanisms in-depth and their impact on the development of kidney disease.</p><p>In comparison, lncRNA was found to regulate gene expression via interacting with RNA molecules, modulating protein post-translational modifications, or remodelling chromatin states despite lacking an open reading framework. m<sup>6</sup>A modification participates in lncRNA biogenesis and regulates numerous kidney cell activities. In glomerulonephritis kidney tissues, METTL3 was increased. To profile the landscape of lncRNAs in glomerular cells, mouse mesangial cells were stimulated with lipopolysaccharide, and then subjected to methylated RNA immunoprecipitation sequencing which identified 1141 differential m<sup>6</sup>A-methylated lncRNAs. These lncRNA-targeted mRNAs were mainly enriched in NF-KB, MAPK, and Toll-like receptor signaling pathways. Knockdown of METTL3 in mesangial cells drastically reduced the levels of m<sup>6</sup>A RNA methylation, cytokines interleukin 6 and tumour necrosis factor-α, and inhibited cell proliferation and cell cycle progression. These results laid a basis for further understanding lncRNA m<sup>6</sup>A modification in the pathogenesis of glomerulonephritis.<span><sup>7</sup></span> Besides glomerular diseases, m<sup>6</sup>A-induced lncRNA MALAT1 aggravated kidney fibrosis by activating the miR-145/FAK axis after obstructive nephropathy.<span><sup>8</sup></span> Impressively, demethylase FTO promoted the epithelial-mesenchymal transition of the cultured tubular cells by reducing the m<sup>6</sup>A modification of lncRNA GAS5. In addition, although lncRNA signature has been used for early diagnosis and prognosis prediction, the role of m<sup>6</sup>A-modified lncRNAs in the occurrence of kidney carcinoma remains unclear.</p><p>As mentioned above, m<sup>6</sup>A modification and ncRNAs are mutually regulated. Mechanistically, most miRNAs modulate m<sup>6</sup>A modification by binding the 3′UTR of mRNAs of m<sup>6</sup>A-related enzymes and inhibiting their translation. Unfortunately, the current evidence of miRNAs regulating m<sup>6</sup>A modification in non-tumor kidney diseases is not robust. Among existing miRNAs, miR-155 is a potential new biomarker for monitoring the progression of lupus nephritis (LN) and may impact the occurrence and development of LN by targeting m<sup>6</sup>A regulators.<span><sup>9</sup></span> In kidney clear cell carcinoma, miR-155 negatively regulated 3′ UTR of mRNA of FTO, an eraser of m<sup>6</sup>A, which increased m<sup>6</sup>A level, reduced apoptosis, and enhanced tumour cell proliferation.<span><sup>10</sup></span> Similarly, miR-501-3p was also reported to inhibit kidney cancer cell proliferation by targeting WTAP and decreasing the level of m<sup>6</sup>A.<span><sup>11</sup></span> Different from miRNAs, lncRNAs often serve as scaffolds and regulate target molecule stability. lncRNA scaffolds with specific binding domains can recruit m<sup>6</sup>A writers, enhancing the m<sup>6</sup>A level of downstream mRNA. Amid kidney fibrosis progression, lncRNA AI662270 binds to the CTGF promoter and directly interacts with METTL3 to increase CTGF mRNA m<sup>6</sup>A methylation and consequently enhance the stability of CTGF mRNA.<span><sup>12</sup></span> Certainly, lncRNA also possesses the ability to determine the prognosis of kidney cell carcinoma, such as stimulating m<sup>6</sup>A in PARP1 mRNA and accelerating the decay of PARP1.<span><sup>13</sup></span></p><p>In summary, we herein showed how m<sup>6</sup>A modification and noncoding RNAs are mutually regulated in kidney diseases. Further understanding the interplay between m<sup>6</sup>A modification and ncRNAs would undoubtedly provide novel perspectives for comprehending the intricate regulatory networks of kidney diseases. As refractory syndromes, kidney diseases often lead to inevitable and irreversible detrimental outcomes in the clinic. Given that m<sup>6</sup>A is a dynamic and reversible process and noncoding RNAs have provoked interest in various diseased scenarios, we expect a bright future of halting kidney disease by explicitly modulating the communications between m<sup>6</sup>A and noncoding RNAs, although it might be a long journey.</p><p>The authors declare no conflict of interest.</p>","PeriodicalId":72605,"journal":{"name":"Clinical and translational discovery","volume":"3 5","pages":""},"PeriodicalIF":0.0000,"publicationDate":"2023-09-29","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1002/ctd2.246","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"Clinical and translational discovery","FirstCategoryId":"1085","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1002/ctd2.246","RegionNum":0,"RegionCategory":null,"ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"","JCRName":"","Score":null,"Total":0}
引用次数: 0
Abstract
First discovered in 1974,1 N6-methyladenosine (m6A) is the most prevalent epigenetic modification in eukaryotic mRNAs at the posttranscriptional level under pathophysiological conditions. The m6A modification entails methylation of adenine at the nitrogen 6 (N6) position. Three classes of enzymes mediate the process of m6A modification, including RNA methyltransferases (writers), demethylases (erasers), and methylation recognition proteins (readers).2 Writers can form a multicomponent m6A methyltransferase complex (MTC) that catalyzes RNA modification by transferring methyl groups to RNA. In MTC, METTL3, METTL14 and WTAP are core components. Serving as erasers, as the name implies, demethylases (FTO and ALKBH5) are responsible for removing S-adenosyl methionine from adenines in RNA. At the same time, readers (YTHDF, YTHDC and eIF3) can recognize m6A-modified RNAs and bind the m6A modification, subsequently determining the fate of the RNA. Therefore, m6A RNA modification is considered invertible and dynamic. In recent years, the advent of advanced high-throughput sequencing has facilitated the identification of m6A.
In general, m6A presents 0.4%−0.6% of all cellular RNAs, including mRNAs, noncoding RNAs (ncRNAs), and circular RNAs (circRNAs).3 In a recent publication in Clinical and Translational Medicine, Ni et al. reported that METTL3 promoted the m6A modification of Ena/VASP-like (EVL) mRNA and subsequently activated the transforming growth factor-beta 1 (TGF-β1) signalling pathway, which ultimately aggravated kidney fibrosis.4 Beyond regulating mRNA-coding genes, m6A modification exhibited remarkable capacity in modulating the function of ncRNAs to dictate kidney fate after various insults, especially in kidney carcinoma. Conversely, ncRNAs also participate in the regulation of m6A modification amid kidney disease progression. Therefore, fully understanding the crosstalk between m6A modification and ncRNAs will supplement a piece of the puzzle of the pathogenesis of non-tumour kidney diseases and shed light on developing effective therapeutic strategies for these medical catastrophes.
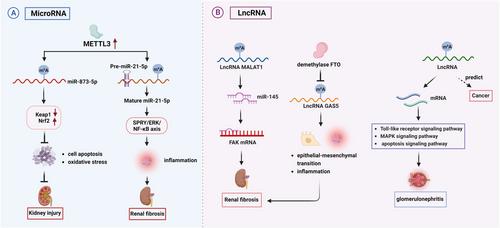
ncRNAs, mainly comprised of micro RNAs (miRNAs), long noncoding RNAs (lncRNAs), and circRNAs, account for a large class of transcripts in mammalian genomes without protein-coding potential. Despite this, they are crucial regulators of diverse physiological and pathological processes. The m6A modification of ncRNAs controls RNA stability, processing, and transport. As illustrated in Figure 1, accumulating studies have shown that m6A-modified ncRNAs cause kidney disorders through multiple dysregulated signalling pathways in diseased kidneys. Because the evidence of m6A modification of circRNAs remains lacking, we mainly focus on introducing the role of miRNAs and lncRNAs in the kidney system. miRNAs make up the majority of ncRNAs, and the role of m6A modification of miRNA is complex and diverse. For instance, in vitro, METTL3 positively modulated miR-873-5p to attenuate oxidative stress and apoptosis by regulating the Keap1/Nrf2 pathway in the cultured proximal tubular cells stimulated with colistin.5 In vivo, METTL3-mediated m6A modification promoted miR-21-5p maturation. Matured miR-21-5p subsequently activated the SPRY1/ERK/NF-κB axis in the obstructive kidneys, exacerbating inflammation and ultimately driving kidney fibrosis.6 As depicted in these findings, m6A modification may exert opposite regulatory roles at different stages of miRNA maturation. Therefore, obtaining a full image of m6A modification of miRNA is required to elucidate the regulatory mechanisms in-depth and their impact on the development of kidney disease.
In comparison, lncRNA was found to regulate gene expression via interacting with RNA molecules, modulating protein post-translational modifications, or remodelling chromatin states despite lacking an open reading framework. m6A modification participates in lncRNA biogenesis and regulates numerous kidney cell activities. In glomerulonephritis kidney tissues, METTL3 was increased. To profile the landscape of lncRNAs in glomerular cells, mouse mesangial cells were stimulated with lipopolysaccharide, and then subjected to methylated RNA immunoprecipitation sequencing which identified 1141 differential m6A-methylated lncRNAs. These lncRNA-targeted mRNAs were mainly enriched in NF-KB, MAPK, and Toll-like receptor signaling pathways. Knockdown of METTL3 in mesangial cells drastically reduced the levels of m6A RNA methylation, cytokines interleukin 6 and tumour necrosis factor-α, and inhibited cell proliferation and cell cycle progression. These results laid a basis for further understanding lncRNA m6A modification in the pathogenesis of glomerulonephritis.7 Besides glomerular diseases, m6A-induced lncRNA MALAT1 aggravated kidney fibrosis by activating the miR-145/FAK axis after obstructive nephropathy.8 Impressively, demethylase FTO promoted the epithelial-mesenchymal transition of the cultured tubular cells by reducing the m6A modification of lncRNA GAS5. In addition, although lncRNA signature has been used for early diagnosis and prognosis prediction, the role of m6A-modified lncRNAs in the occurrence of kidney carcinoma remains unclear.
As mentioned above, m6A modification and ncRNAs are mutually regulated. Mechanistically, most miRNAs modulate m6A modification by binding the 3′UTR of mRNAs of m6A-related enzymes and inhibiting their translation. Unfortunately, the current evidence of miRNAs regulating m6A modification in non-tumor kidney diseases is not robust. Among existing miRNAs, miR-155 is a potential new biomarker for monitoring the progression of lupus nephritis (LN) and may impact the occurrence and development of LN by targeting m6A regulators.9 In kidney clear cell carcinoma, miR-155 negatively regulated 3′ UTR of mRNA of FTO, an eraser of m6A, which increased m6A level, reduced apoptosis, and enhanced tumour cell proliferation.10 Similarly, miR-501-3p was also reported to inhibit kidney cancer cell proliferation by targeting WTAP and decreasing the level of m6A.11 Different from miRNAs, lncRNAs often serve as scaffolds and regulate target molecule stability. lncRNA scaffolds with specific binding domains can recruit m6A writers, enhancing the m6A level of downstream mRNA. Amid kidney fibrosis progression, lncRNA AI662270 binds to the CTGF promoter and directly interacts with METTL3 to increase CTGF mRNA m6A methylation and consequently enhance the stability of CTGF mRNA.12 Certainly, lncRNA also possesses the ability to determine the prognosis of kidney cell carcinoma, such as stimulating m6A in PARP1 mRNA and accelerating the decay of PARP1.13
In summary, we herein showed how m6A modification and noncoding RNAs are mutually regulated in kidney diseases. Further understanding the interplay between m6A modification and ncRNAs would undoubtedly provide novel perspectives for comprehending the intricate regulatory networks of kidney diseases. As refractory syndromes, kidney diseases often lead to inevitable and irreversible detrimental outcomes in the clinic. Given that m6A is a dynamic and reversible process and noncoding RNAs have provoked interest in various diseased scenarios, we expect a bright future of halting kidney disease by explicitly modulating the communications between m6A and noncoding RNAs, although it might be a long journey.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


