{"title":"‘Splicing predictions, minigene analyses, and ACMG-AMP clinical classification of 42 germline PALB2 splice-site variants’","authors":"","doi":"10.1002/path.6215","DOIUrl":null,"url":null,"abstract":"<p>By Alberto Valenzuela-Palomo, Elena Bueno-Martínez, Lara Sanoguera-Miralles, Víctor Lorca, Eugenia Fraile-Bethencourt, Ada Esteban-Sánchez, Susana Gómez-Barrero, Sara Carvalho, Jamie Allen, Alicia García-Álvarez, Pedro Pérez-Segura, Leila Dorling, Douglas F Easton, Peter Devilee, Maaike PG Vreeswijk, Miguel de la Hoya<sup>†</sup> and Eladio A Velasco<sup>†</sup>, <i>J Pathol</i> 2022; <b>256:</b> 321–334. https://doi.org/10.1002/path.5839. <sup>†</sup>Senior authors.</p><p>The senior authors of this article, first published on 30 November 2021 in Wiley Online Library (wileyonlinelibrary.com), have informed the editors of an unintentional mistake that they wish to correct, and they explain why below.</p><p>In our manuscript, we described minigene splicing read-outs for 42 <i>PALB2</i> variants. Two of them, c.1685-2A>C and c.1685-2A>G (acceptor site of exon 5), displayed a complex pattern of five anomalous transcripts: ∆(E5p139), ∆(E5p5), ▼(E5p88), ∆(E5p10) and ∆(E5p97) (Table 1 and Figure 2B of the original manuscript).</p><p>Therefore, Δ(E5p510) represents roughly 18% and 25% of the overall minigene expression of c.1685-2A>C and c.1685-2A>G, respectively.</p><p>In the light of our new data, we re-evaluated the ACMG-AMP point-based system classification of both variants.</p><p>Since all the initially minigene-detected transcripts were PTC_NMD (∆(E5p139), ▼(E5p88), ∆(E5p10), ∆(E5p97) and ∆(E5p5)), we had previously assigned PS3_VS (+8) to both variants (supplementary material, Table S2, and Table 2). Yet, ∆(E5p510) makes a substantial contribution to the overall expression. Moreover, this transcript encodes for an in-frame protein deletion, p.(Lys563_Gly732del), which removes >10% of the protein sequence but does not target known critical PALB2 domains, so that ∆(E5p510) finally qualifies for a PS3 code.</p><p><b>Figure C1.</b> Fluorescent fragment analysis of the wild type minigene mgPALB2_ex4-6 and variants c.1685-2A>C and c.1685-2A>G. FAM-labeled products (blue peaks) were run with LIZ-1200 (orange peaks) as size standards. The <i>x</i>-axis indicates size in bp (electropherogram on the top) and the <i>y</i>-axis represents relative fluorescence units (RFU). FL, minigene full-length transcript. As indicated in Materials and methods, cDNA was amplified with primers RTPB2_EX4-FW (5’-CACAAATATCAGCACGAAAA-3’) and FAM-RTPB2_EX6-RV (full-length size: 918 nt).</p><p>The authors apologise for any inconvenience this mistake may have caused.</p><p>The editors apologise for the time taken to process the request for this corrigendum and approve it.</p>","PeriodicalId":232,"journal":{"name":"The Journal of Pathology","volume":"261 3","pages":"372-373"},"PeriodicalIF":5.6000,"publicationDate":"2023-09-27","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1002/path.6215","citationCount":"1","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"The Journal of Pathology","FirstCategoryId":"3","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1002/path.6215","RegionNum":2,"RegionCategory":"医学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"ONCOLOGY","Score":null,"Total":0}
引用次数: 1
Abstract
By Alberto Valenzuela-Palomo, Elena Bueno-Martínez, Lara Sanoguera-Miralles, Víctor Lorca, Eugenia Fraile-Bethencourt, Ada Esteban-Sánchez, Susana Gómez-Barrero, Sara Carvalho, Jamie Allen, Alicia García-Álvarez, Pedro Pérez-Segura, Leila Dorling, Douglas F Easton, Peter Devilee, Maaike PG Vreeswijk, Miguel de la Hoya† and Eladio A Velasco†, J Pathol 2022; 256: 321–334. https://doi.org/10.1002/path.5839. †Senior authors.
The senior authors of this article, first published on 30 November 2021 in Wiley Online Library (wileyonlinelibrary.com), have informed the editors of an unintentional mistake that they wish to correct, and they explain why below.
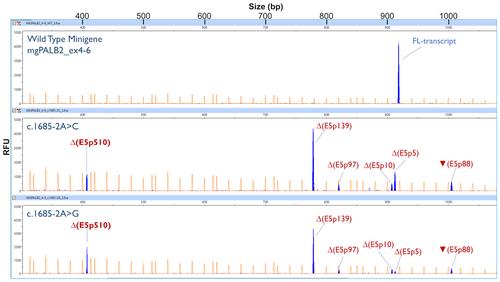
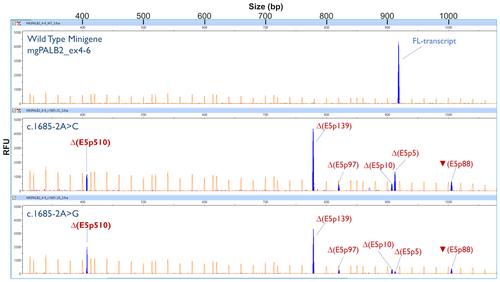
In our manuscript, we described minigene splicing read-outs for 42 PALB2 variants. Two of them, c.1685-2A>C and c.1685-2A>G (acceptor site of exon 5), displayed a complex pattern of five anomalous transcripts: ∆(E5p139), ∆(E5p5), ▼(E5p88), ∆(E5p10) and ∆(E5p97) (Table 1 and Figure 2B of the original manuscript).
Therefore, Δ(E5p510) represents roughly 18% and 25% of the overall minigene expression of c.1685-2A>C and c.1685-2A>G, respectively.
In the light of our new data, we re-evaluated the ACMG-AMP point-based system classification of both variants.
Since all the initially minigene-detected transcripts were PTC_NMD (∆(E5p139), ▼(E5p88), ∆(E5p10), ∆(E5p97) and ∆(E5p5)), we had previously assigned PS3_VS (+8) to both variants (supplementary material, Table S2, and Table 2). Yet, ∆(E5p510) makes a substantial contribution to the overall expression. Moreover, this transcript encodes for an in-frame protein deletion, p.(Lys563_Gly732del), which removes >10% of the protein sequence but does not target known critical PALB2 domains, so that ∆(E5p510) finally qualifies for a PS3 code.
Figure C1. Fluorescent fragment analysis of the wild type minigene mgPALB2_ex4-6 and variants c.1685-2A>C and c.1685-2A>G. FAM-labeled products (blue peaks) were run with LIZ-1200 (orange peaks) as size standards. The x-axis indicates size in bp (electropherogram on the top) and the y-axis represents relative fluorescence units (RFU). FL, minigene full-length transcript. As indicated in Materials and methods, cDNA was amplified with primers RTPB2_EX4-FW (5’-CACAAATATCAGCACGAAAA-3’) and FAM-RTPB2_EX6-RV (full-length size: 918 nt).
The authors apologise for any inconvenience this mistake may have caused.
The editors apologise for the time taken to process the request for this corrigendum and approve it.
期刊介绍:
The Journal of Pathology aims to serve as a translational bridge between basic biomedical science and clinical medicine with particular emphasis on, but not restricted to, tissue based studies. The main interests of the Journal lie in publishing studies that further our understanding the pathophysiological and pathogenetic mechanisms of human disease.
The Journal of Pathology welcomes investigative studies on human tissues, in vitro and in vivo experimental studies, and investigations based on animal models with a clear relevance to human disease, including transgenic systems.
As well as original research papers, the Journal seeks to provide rapid publication in a variety of other formats, including editorials, review articles, commentaries and perspectives and other features, both contributed and solicited.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


